
Resource Screening and Inheritance Analysis of Fusarium oxysporum sp. conglutinans Race 2 Resistance in Cabbage (Brassica oleracea var. capitata)

 and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Materials
2.2. Inoculation and Resistance Testing
2.3. Data Collection and Analysis
2.4. Genetic Analyses
3. Results
3.1. Screening for Race 1 and 2 CFW Resistance
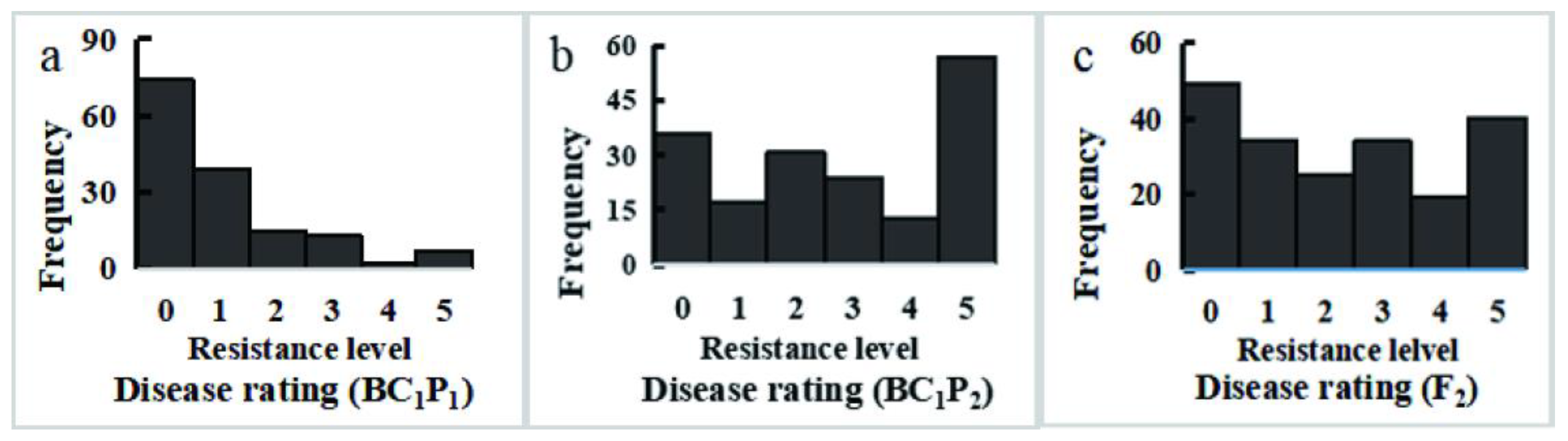
3.2. CFW Resistance Frequency Distributions among Segregating Populations
3.3. Optimal Genetic Model Selection and Testing
3.4. Genetic Parameter Estimations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Smith, E. The fungus infection of agricultural soils in the United States. Sci. Am. Suppl. 1899, 48, 19981–19982. [Google Scholar]
- Nomura, Y. Studies on the method of early selection of the resistance of cabbage to the yellows disease. J. Cent. Agric. Exp. Stn. 1976, 24, 141–182. [Google Scholar]
- Thanassoulopoulos, C.; Kitsos, G.; Bonatsos, D. Cabbage yellows and new hosts of the pathogen in Greece. Plant Dis. Report. 1978, 62, 1051–1053. [Google Scholar]
- Howell, E.; Barker, G.; Jones, G.; Kearsey, M.; King, G.; Kop, E.; Armstrong, S. Integration of the cytogenetic and genetic linkage maps of Brassica oleracea. Genetics 2002, 161, 1225–1234. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, T.; Li, X.; Yan, H. Fusarium wilt of cruciferae and its pathogen identifification. Plant Prot. 2003, 29, 44–45. (In Chinese) [Google Scholar]
- Gilman, J. Cabbage yellows and the relation of temperature to its occurrence. Ann. Mo. Bot. Gard. 1916, 3, 25–84. [Google Scholar] [CrossRef]
- Ramirez Villupadua, J.; Endo, R.; Bosland, P.; Wiliams, P. A new race of Fusarium oxysporum f. sp. conglutinans that attacks cabbage with type A resistance. Plant Dis. 1985, 69, 612–613. [Google Scholar]
- Bosland, P.; Williams, P. Pathogenicity of geographic isolates of Fusarium oxysporum from crucifers on a differential set of crucifer seedlings. J. Phytopathol. 1988, 123, 63–68. [Google Scholar] [CrossRef]
- Jones, L.; Walker, J.; Tisdale, W. Fusarium resistant cabbage. Agric. Exp. Stn. Univ. Wis. 1920, 48, 34. [Google Scholar]
- Monteiro, A.; Williams, P. The exploration of genetic resources of Portuguese cabbage and kale for resistance to several Brassica diseases. Euphytica 1989, 41, 215–225. [Google Scholar] [CrossRef]
- Wang, J.; Podlich, D.; Cooper, M.; DeLacy, I. Power of the joint segregation analysis method for testing mixed major-gene and polygene inheritance models of quantitative traits. Theor. Appl. Genet. 2001, 103, 804–816. [Google Scholar] [CrossRef]
- Qi, Z.; Li, J.; Raza, M.; Zou, X.; Cao, L.; Rao, L.; Chen, L. Inheritance of fruit cracking resistance of melon (Cucumis melo L.) fitting E-0 genetic model using major gene plus polygene inheritance analysis. Sci. Hortic. 2015, 189, 168–174. [Google Scholar] [CrossRef]
- Sun, X.; Liu, L.; Zhi, X.; Bai, J.; Cui, Y.; Shu, J.; Li, J. Genetic analysis of tomato internode length via mixed major gene plus polygene inheritance model. Sci. Hortic. 2019, 246, 759–764. [Google Scholar] [CrossRef]
- Kong, C.; Chen, G.; Yang, L.; Zhuang, M.; Zhang, Y.; Wang, Y.; Ji, J.; Fang, Z.; Lv, H. Germplasm screening and inheritance analysis of resistance to cabbage black rot in a worldwide collection of cabbage (Brassica oleracea var. capitata) resources. Sci. Hortic. 2021, 288, 110234. [Google Scholar] [CrossRef]
- Lv, H.; Fang, Z.; Yang, L.; Zhang, Y.; Wang, Q.; Liu, Y.; Zhuang, M.; Yang, Y.; Xie, B.; Liu, B.; et al. Mapping and analysis of a novel candidate Fusarium wilt resistance gene FOC1 in Brassica Oleracea. BMC Genom. 2014, 15, 1094. [Google Scholar] [CrossRef]
- Shimizu, M.; Fujimoto, R.; Ying, H.; Pu, Z.; Ebe, Y.; Kawanabe; Saeki, N.; Taylor, J.; Kaji, M.; Dennis, E.; et al. Identification of candidate genes for Fusarium yellows resistance in Chinese cabbage by differential expression analysis. Plant Mol. Biol. 2014, 85, 247–257. [Google Scholar] [CrossRef]
- Shimizu, M.; Pu, Z.; Kawanabe, T.; Kitashiba, H.; Matsumoto, S.; Ebe, Y.; Sano, M.; Funaki, T.; Fukai, E.; Fujimoto, R.; et al. Map-based cloning of a candidate gene conferring Fusarium yellows resistance in Brassica Oleracea. Theor. Appl. Genet. 2015, 128, 119–130. [Google Scholar] [CrossRef]
- Blank, L. The pathogenicity of Fusarium conglutinans Wr. at low soil temperatures. Phytopathology 1932, 22, 191–195. [Google Scholar]
- Blank, L. Fusarium resistance in Wisconsin all seasons cabbage. J. Agric. Res. 1937, 55, 497–510. [Google Scholar]
- Walker, J.; Hooker, W. Plant nutrition in relation to disease development. I. Cabbage yellows. Am. J. Bot. 1945, 32, 314–320. [Google Scholar] [CrossRef]
- Bosland, P.; Williams, P. An evaluation of Fusarium oxysporum from crucifers based on pathogenicity, isozyme polymorphism, vegetative compatibility, and geographic origin. Can. J. Bot. 1987, 65, 2067–2073. [Google Scholar] [CrossRef]
- Bosland, P.; Williams, P. Sources of resistance to Fusarium oxysporum f. sp. conglutinans, race 2. HortScience 1987, 22, 669–670. [Google Scholar]
- Liu, X.; Ling, J.; Xiao, Z.; Xie, B.; Fang, Z.; Yang, L.; Yang, Y. Characterization of emerging populations of Fusarium oxysporum f. sp. conglutinans causing cabbage wilt in China. J. Phytopathol. 2017, 165, 813–821. [Google Scholar]
- Liu, Z.; Xie, J.; Wang, H.; Zhong, X.; Li, H.; Yu, J.; Kang, J. Identification and expression profiling analysis of NBS-LRR genes involved in Fusarium oxysporum f. sp. conglutinans resistance in cabbage. 3 Biotech 2019, 9, 202. [Google Scholar] [PubMed]
- Lv, H.; Wang, Q.; Liu, X.; Han, F.; Fang, Z.; Yang, L.; Zhang, Y. Whole-genome mapping reveals novel QTL clusters associated with main agronomic traits of cabbage (Brassica oleracea var. capitata L.). Front. Plant Sci. 2016, 7, 989. [Google Scholar] [CrossRef]
- Lv, H.; Yang, L.; Kang, J.; Wang, Q.; Wang, X.; Fang, Z.; Liu, J. Development of InDel markers linked to Fusarium wilt resistance in cabbage. Mol. Breed. 2013, 32, 961–967. [Google Scholar] [CrossRef]
- Liu, X.; Han, F.; Kong, C.; Fang, Z.; Yang, L.; Zhang, Y.; Zhuang, M.; Liu, Y.; Li, Z.; Lv, H. Rapid introgression of the Fusarium wilt resistance gene into an elite cabbage line through the combined application of a microspore culture, genome background analysis, and disease resistance-specific marker assisted foreground selection. Front. Plant Sci. 2017, 8, 354. [Google Scholar] [CrossRef]
- Liu, X.; Xing, M.; Kong, C.; Fang, Z.; Yang, L.; Zhang, Y.; Wang, Y.; Ling, J.; Yang, Y.; Lv, H. Genetic diversity, virulence, race profiling, and comparative genomic analysis of the Fusarium oxysporum f. sp. conglutinans strains infecting cabbages in China. Front. Microbiol. 2019, 10, 1373. [Google Scholar]
- Gai, J.; Zhang, Y.; Wang, J. Genetic System of Quantitative Traits in Plants; Science Press: Beijing, China, 2003; Volume 8, pp. 224–260. (In Chinese) [Google Scholar]
- Booth, C. The Genus Fusarium; Commonwealth Mycological Institute: Kew Surrey, UK, 1971; 237p. [Google Scholar]
- Lv, H.; Fang, Z.; Yang, L.; Zhang, Y.; Wang, Y. An update on the arsena l: Mining resistance genes for disease management of Brassica crops in the genomic era. Hortic. Res. 2020, 7, 34. [Google Scholar] [CrossRef]
- Diaz, J.; Garcia, J.; Lara, C.; Hutmacher, R.; Ulloa, M.; Nichols, R.; Ellis, M. Characterization of current Fusarium oxysporum f. sp. vasinfectum isolates from cotton in the San Joaquin Valley of California and Lower Valley El Paso, Texas. Plant Dis. 2021, 105, 1898–1911. [Google Scholar]
- Amaradasa, B.; Beckham, K.; Dufault, N.; Sanchez, T.; Ertek, T.; Iriarte, F.; Ji, P. First report of Fusarium oxysporum f. sp. niveum race 3 causing wilt of watermelon in Florida, USA. Plant Dis. 2018, 102, 1029. [Google Scholar]
- Anderson, J.; Aitken, E. Effect of in planta treatment of ‘Cavendish’ banana with herbicides and fungicides on the colonisation and sporulation by Fusarium oxysporum f. sp. cubense subtropical race 4. J. Fungi 2021, 7, 184. [Google Scholar]
- Paulino, J.; Almeida, C.; Gonçalves, G.; Bueno, C.; Carbonell, S.; Chiorato, A.; Bechimol-Reis, L. Assessment of resistance in common bean to Fusarium oxysporum f. sp. phaseoli using different inoculation and evaluation methods. Crop Breed. Appl. Biotechnol. 2020, 20, 2020. [Google Scholar]
- Morrison, R.; Mengistu, A.; Williams, P. First report of race 2 of cabbage yellows caused by Fusarium oxysporum f. sp. conglutinans in Texas. Plant Dis. 1994, 78, 641. [Google Scholar] [CrossRef]
- Walker, J. Inheritance of Fusarium resistance in cabbage. J. Agric. Res. 1930, 40, 721–745. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Race 1 | Race 2 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Number of Accessions | Percentage of Plants (%) | Number of Accessions | Percentage of Plants (%) | ||||||||||
| Classification | Highly Resistant | Resistant | Moderately Resistant | Susceptible | Highly Susceptible | Highly Resistant | Resistant | Moderately Resistant | Susceptible | Highly Susceptible | |||
| Geographic origin | China | 18 | 5.56 | 22.22 | 27.78 | 16.67 | 27.78 | 18 | 5.56 | 5.56 | 11.11 | 22.22 | 55.56 |
| Asia except China | 107 | 42.06 | 13.08 | 17.76 | 14.95 | 12.15 | 107 | 35.51 | 12.15 | 11.21 | 12.15 | 28.97 | |
| North America | 12 | 41.67 | 8.33 | 16.67 | 8.33 | 25.00 | 12 | 33.33 | 8.33 | 8.33 | 16.67 | 33.33 | |
| Europe | 29 | 20.69 | 6.90 | 20.69 | 27.59 | 24.14 | 29 | 6.90 | 13.79 | 20.69 | 10.34 | 48.28 | |
| Planting season | Spring | 43 | 23.26 | 4.65 | 27.91 | 20.93 | 23.26 | 43 | 11.63 | 6.98 | 13.95 | 20.93 | 46.51 |
| Autumn | 90 | 40.00 | 14.44 | 15.56 | 14.44 | 15.56 | 90 | 32.22 | 11.11 | 12.22 | 12.22 | 32.22 | |
| Overwintering | 33 | 33.33 | 18.18 | 18.18 | 18.18 | 12.12 | 33 | 33.33 | 18.18 | 12.12 | 6.06 | 30.30 | |
| Maturity | Early maturity | 53 | 39.62 | 7.55 | 20.75 | 13.21 | 18.87 | 53 | 24.53 | 11.32 | 11.32 | 11.32 | 41.51 |
| Mid-early maturity | 38 | 34.21 | 10.53 | 18.42 | 18.42 | 18.42 | 38 | 28.95 | 10.53 | 13.16 | 18.42 | 28.95 | |
| Medium maturity | 9 | 55.56 | 11.11 | 22.22 | 11.11 | 0.00 | 9 | 33.33 | 33.33 | 11.11 | 11.11 | 11.11 | |
| Mid-late maturity | 12 | 25.00 | 0.00 | 25.00 | 25.00 | 25.00 | 12 | 25.00 | 0.00 | 16.67 | 16.67 | 41.67 | |
| Late maturity | 54 | 27.78 | 22.22 | 16.67 | 20.37 | 12.96 | 54 | 27.78 | 11.11 | 12.96 | 11.11 | 37.04 | |
| Head shape | Flat | 59 | 28.81 | 18.64 | 22.03 | 16.95 | 13.56 | 59 | 28.81 | 11.86 | 13.56 | 13.56 | 32.20 |
| Round | 107 | 37.38 | 9.35 | 17.76 | 17.76 | 17.76 | 107 | 26.17 | 11.21 | 12.15 | 13.08 | 37.38 | |
| Leaf color | Grey | 7 | 71.43 | 0.00 | 0.00 | 0.00 | 28.57 | 7 | 57.14 | 14.29 | 0.00 | 0.00 | 28.57 |
| Gray green | 67 | 33.33 | 15.79 | 21.05 | 17.54 | 12.28 | 67 | 29.82 | 12.28 | 12.28 | 33.33 | 12.28 | |
| Green | 64 | 28.13 | 17.19 | 17.19 | 17.19 | 20.31 | 64 | 18.75 | 14.06 | 15.63 | 14.06 | 37.50 | |
| Dark green | 38 | 39.47 | 2.63 | 23.68 | 18.42 | 15.79 | 38 | 31.58 | 5.26 | 10.53 | 10.53 | 42.11 | |
| Generation | Number | Frequency Distribution of FW Disease Rating in Each Population | Mean Disease Index | |||||
|---|---|---|---|---|---|---|---|---|
| Level 0 | Level 1 | Level 2 | Level 3 | Level 4 | Level 5 | |||
| P1 (BI-16) | 15 | 15 | 0 | 0 | 0 | 0 | 0 | 0.00 |
| P2 (01-20) | 15 | 0 | 0 | 0 | 1 | 6 | 8 | 92.00 |
| F1 (BI-16 × 01-20) | 22 | 4 | 6 | 5 | 6 | 0 | 1 | 35.45 |
| BC1P1 (BI-16 × 01-20 × BI-16) | 150 | 74 | 39 | 15 | 13 | 2 | 7 | 20.13 |
| BC1P2 (BI-16 × 01-20 × 01-20) | 178 | 36 | 17 | 31 | 24 | 13 | 57 | 54.83 |
| F2 (BI-16 × 01-20) ⊗ | 201 | 49 | 34 | 25 | 34 | 19 | 40 | 45.97 |
| Model | Implication of Model | Maximum Likelihood Value | AIC |
|---|---|---|---|
| A-1 | 1 MG-AD | −1073.03 | 2154.06 |
| A-2 | 1 MG-A | −1110.42 | 2226.84 |
| A-3 | 1 MG-EAD | −1082.03 | 2170.05 |
| A-4 | 1 MG-AEND | −1156.68 | 2319.36 |
| B-1 | 2MG-AD1 | −1023.43 | 2066.86 |
| B-2 | 2MG-AD | −1070.37 | 2152.73 |
| B-3 | 2MG-A | −1165.04 | 2338.08 |
| B-4 | 2MG-EA | −1123.04 | 2252.09 |
| B-5 | 2MG-AED | −1081.75 | 2171.50 |
| B-6 | 2MG-EEAD | −1105.63 | 2217.25 |
| C | PG-ADI | −1101.32 | 2222.63 |
| C-1 | PG-AD | −1106.42 | 2226.85 |
| D | MX1-AD-ADI | −1103.08 | 2230.17 |
| D-1 | MX1-AD-AD | −1096.19 | 2210.38 |
| D-2 | MX1-A-AD | −1096.19 | 2196.55 |
| D-3 | MX1-EAD-AD | −1081.53 | 2179.06 |
| D-4 | MX1-AEND-AD | −1101.55 | 2219.10 |
| E * | MX2-ADI-ADI | −988.78 | 2013.56 |
| E-1 * | MX2-ADI-AD | −1009.09 | 2048.19 |
| E-2 | MX2-AD-AD | −1055.47 | 2132.94 |
| E-3 * | MX2-A-AD | −1013.12 | 2044.25 |
| E-4 | MX2-EAED-AD | −1106.06 | 2228.11 |
| E-5 | MX2-AED-AD | −1054.71 | 2127.42 |
| Model | Generation | nW2 | Dn | |||
|---|---|---|---|---|---|---|
| E | P1 | 0.00 (1.00) | 1.17 (0.28) | 18.75 (0.00) * | 1.05 (<0.05) * | 0.40 (<0.05) * |
| P2 | 0.05 (0.83) | 1.40 (0.24) | 30.82 (0.00) * | 0.45 (>0.05) | 0.26 (>0.05) | |
| F1 | 0.09 (0.76) | 0.34 (0.56) | 1.35 (0.25) | 0.15 (>0.05) | 0.26 (>0.05) | |
| BC1P1 | 5.03 (0.02) * | 5.12 (0.02) * | 0.13 (0.72) | 2.02 (<0.05) * | 0.21 (<0.05) * | |
| BC1P2 | 3.51 (0.06) | 1.84 (0.17) | 3.33 (0.07) | 1.21 (<0.05) * | 0.08 (>0.05) | |
| F2 | 3.30 (0.07) | 4.03 (0.04) * | 0.99 (0.32) | 0.86 (<0.05) * | 0.67 (>0.05) | |
| E-1 | P1 | 12.95 (0.00) * | 13.19 (0.00) * | 0.35 (0.55) | 2.13 (<0.05) * | 0.67 (<0.05) * |
| P2 | 4.55 (0.03) * | 0.54 (0.46) | 28.27 (0.00) * | 0.79 (<0.05) * | 0.34 (>0.05) | |
| F1 | 1.81 (0.18) | 1.41 (0.23) | 0.21 (0.65) | 0.25 (>0.05) | 0.26 (>0.05) | |
| BC1P1 | 0.21 (0.65) | 0.78 (0.38) | 3.14 (0.08) | 1.62 (<0.05) * | 0.15 (<0.05) * | |
| BC1P2 | 2.54 (0.11) | 1.47 (0.23) | 1.76 (0.19) | 0.99 (<0.05) * | 0.07 (<0.05) * | |
| F2 | 0.15 (0.70) | 0.01 (0.92) | 1.29 (0.26) | 0.47 (>0.05) | 0.03 (<0.05) * | |
| E-3 | P1 | 17.55 (1.00) | 14.99 (0.00) * | 0.54 (0.46) | 2.51 (<0.05) * | 0.71 (<0.05) * |
| P2 | 6.04 (0.01) * | 1.18 (0.28) | 26.75 (0.00) * | 0.89 (<0.05) * | 0.34 (>0.05) | |
| F1 | 2.04 (0.15) | 1.70 (0.19) | 0.11 (0.75) | 0.27 (>0.05) | 0.26 (>0.05) | |
| BC1P1 | 0.09 (0.76) | 0.47 (0.49) | 2.50 (0.11) | 1.59 (<0.05) * | 0.15 (>0.05) | |
| BC1P2 | 2.64 (0.10) | 1.38 (0.24) | 2.57 (0.11) | 0.99 (<0.05) * | 0.07 (<0.05) * | |
| F2 | 6.21 (0.01) * | 5.08 (0.02) * | 0.41 (0.52) | 1.12 (<0.05) * | 0.08 (<0.05) * |
| First-Order Parameter | Estimate | Second-Order Parameter | Estimate | ||
|---|---|---|---|---|---|
| B1 | B2 | F2 | |||
| m1 | 2.08 | σmg2 | 0.58 | 2.67 | 2.40 |
| m2 | 3.00 | σpg2 | 0.89 | 0.66 | 0.66 |
| m3 | 1.74 | σp2 | 1.81 | 3.66 | 3.39 |
| m4 | 2.54 | σe2 | 0.33 | 0.33 | 0.33 |
| m5 | 2.02 | hmg2 | 32.14 | 72.80 | 70.64 |
| m6 | 1.96 | hpg2 | 49.47 | 18.13 | 19.57 |
| da | −1.25 | 1 − (hmg2 + hpg2) | 18.38 | 9.07 | 9.79 |
| db | −1.15 | ||||
| ha | −0.78 | ||||
| hb | 0.29 | ||||
| ha/da | 0.62 | ||||
| hb/db | −0.26 | ||||
| i | 0.32 | ||||
| jab | −1.07 | ||||
| jba | 0.98 | ||||
| l | 2.12 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tong, L.; Zhao, C.; Liu, J.; Yang, L.; Zhuang, M.; Zhang, Y.; Wang, Y.; Ji, J.; Kuang, B.; Tang, K.; et al. Resource Screening and Inheritance Analysis of Fusarium oxysporum sp. conglutinans Race 2 Resistance in Cabbage (Brassica oleracea var. capitata). Genes 2022, 13, 1590. https://doi.org/10.3390/genes13091590
Tong L, Zhao C, Liu J, Yang L, Zhuang M, Zhang Y, Wang Y, Ji J, Kuang B, Tang K, et al. Resource Screening and Inheritance Analysis of Fusarium oxysporum sp. conglutinans Race 2 Resistance in Cabbage (Brassica oleracea var. capitata). Genes. 2022; 13(9):1590. https://doi.org/10.3390/genes13091590
Chicago/Turabian StyleTong, Long, Cunbao Zhao, Jinhui Liu, Limei Yang, Mu Zhuang, Yangyong Zhang, Yong Wang, Jialei Ji, Bifeng Kuang, Kelan Tang, and et al. 2022. "Resource Screening and Inheritance Analysis of Fusarium oxysporum sp. conglutinans Race 2 Resistance in Cabbage (Brassica oleracea var. capitata)" Genes 13, no. 9: 1590. https://doi.org/10.3390/genes13091590