
Pathogenicity of Beauveria bassiana PfBb and Immune Responses of a Non-Target Host, Spodoptera frugiperda (Lepidoptera: Noctuidae)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Entomopathogenic Fungus
2.3. Virulence of B. bassiana PfBb on S. frugiperda Larvae
2.4. Time-Dose-Mortality (TDM) Model of B. bassiana PfBb Affecting S. frugiperda
2.5. Enzymatic Sample Preparation
2.6. Measurement of Enzyme Activities in S. frugiperda Larvae
2.7. Statistical Analyses
3. Results
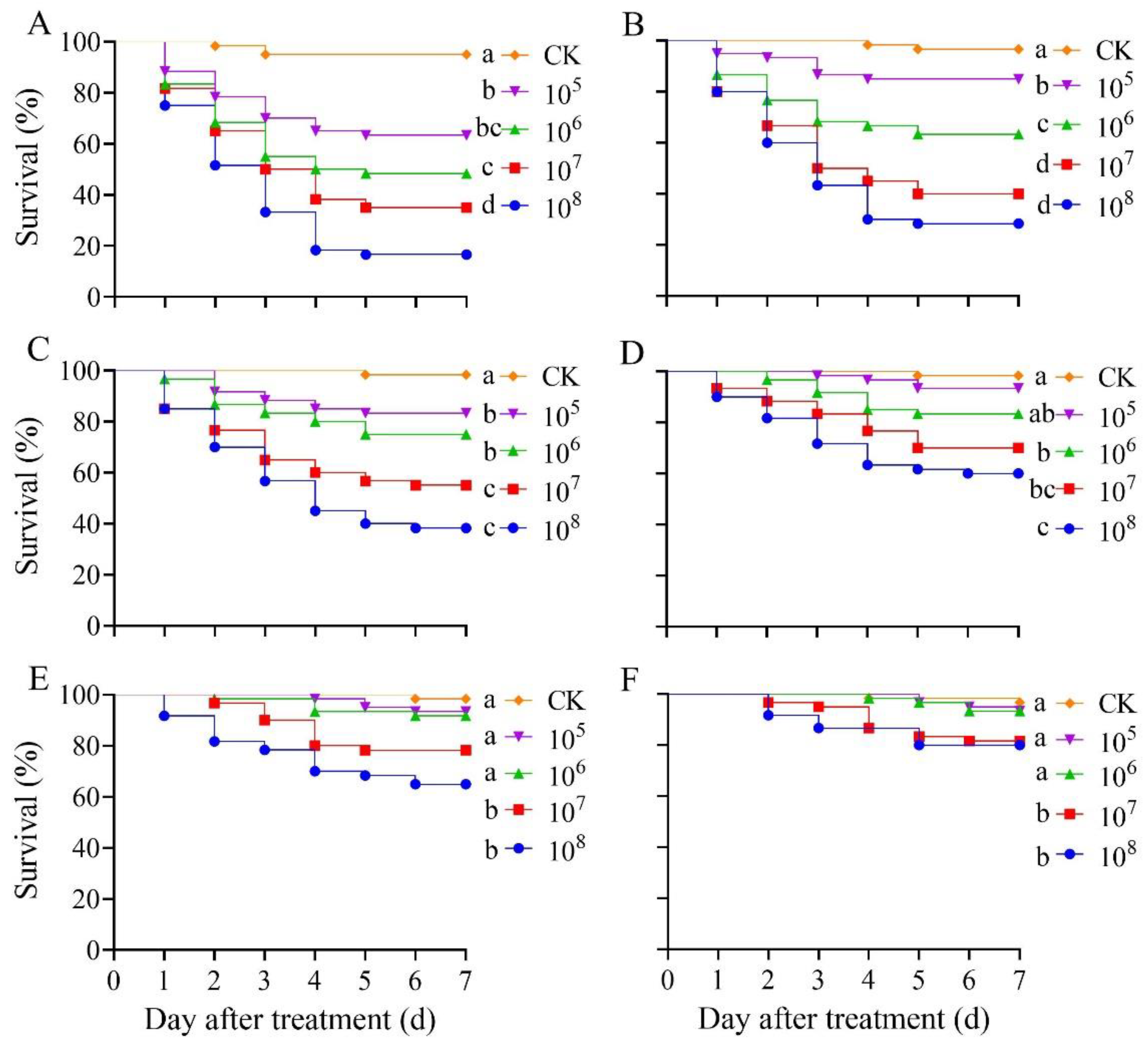
3.1. Effect of B. bassiana PfBb on S. frugiperda Larvae
3.2. Effect of B. bassiana PfBb on Mortality of S. frugiperda Larvae
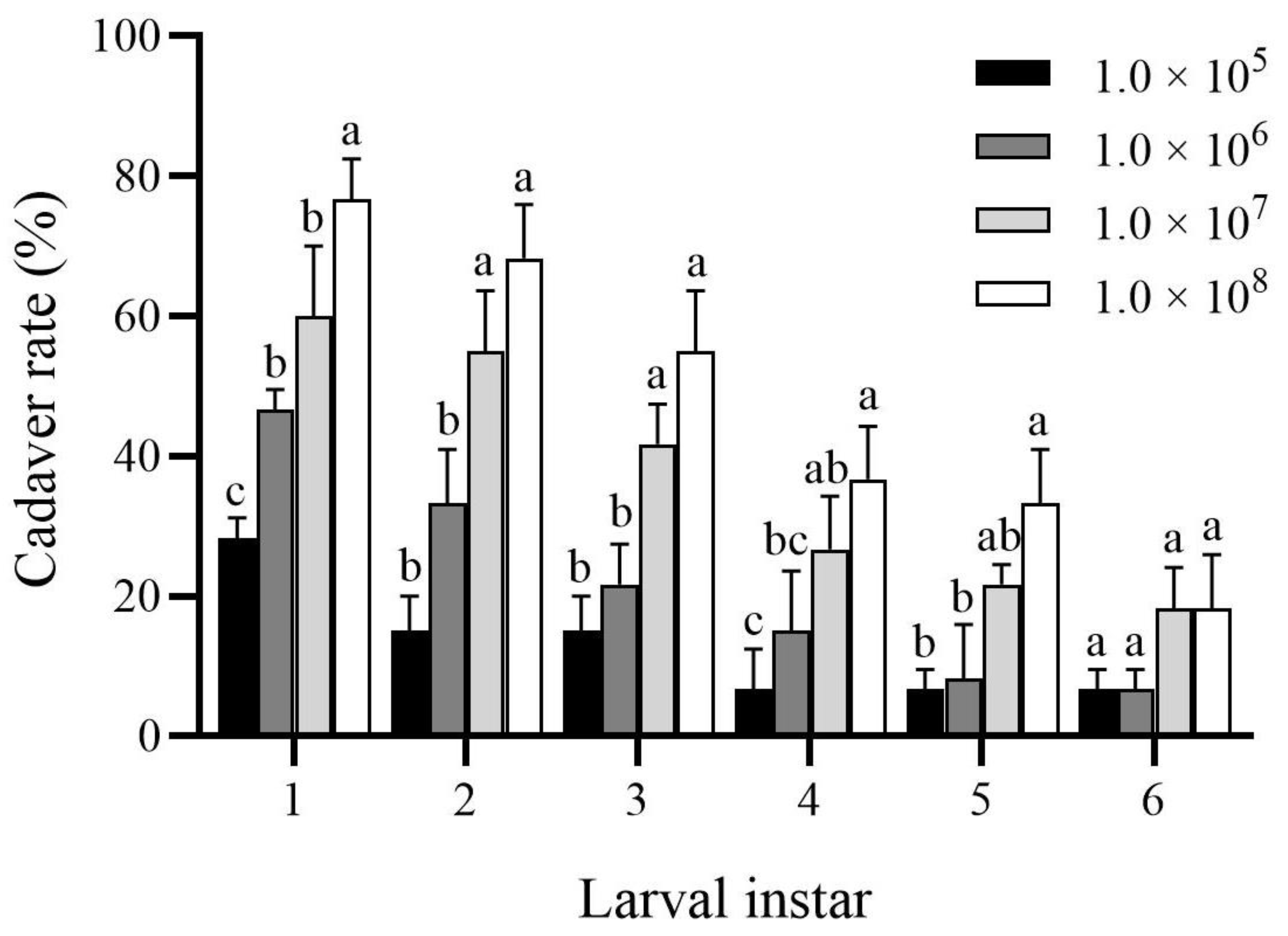
3.3. Effect of B. bassiana PfBb on the Cadaver Rate of Infected S. frugiperda Larvae
3.4. Time-Dose-Mortality (TDM) Model of B. bassiana PfBb on S. frugiperda Larvae
3.5. Effect of B. bassiana PfBb Concentration on the Lethality of S. frugiperda Larvae
3.6. Effect of B. bassiana PfBb on CAT Enzyme Activity in S. frugiperda Larvae
3.7. Effect of B. bassiana PfBb on POD Enzyme Activity in S. frugiperda Larvae
3.8. Effect of B. bassiana PfBb on SOD Enzyme Activity in S. frugiperda Larvae
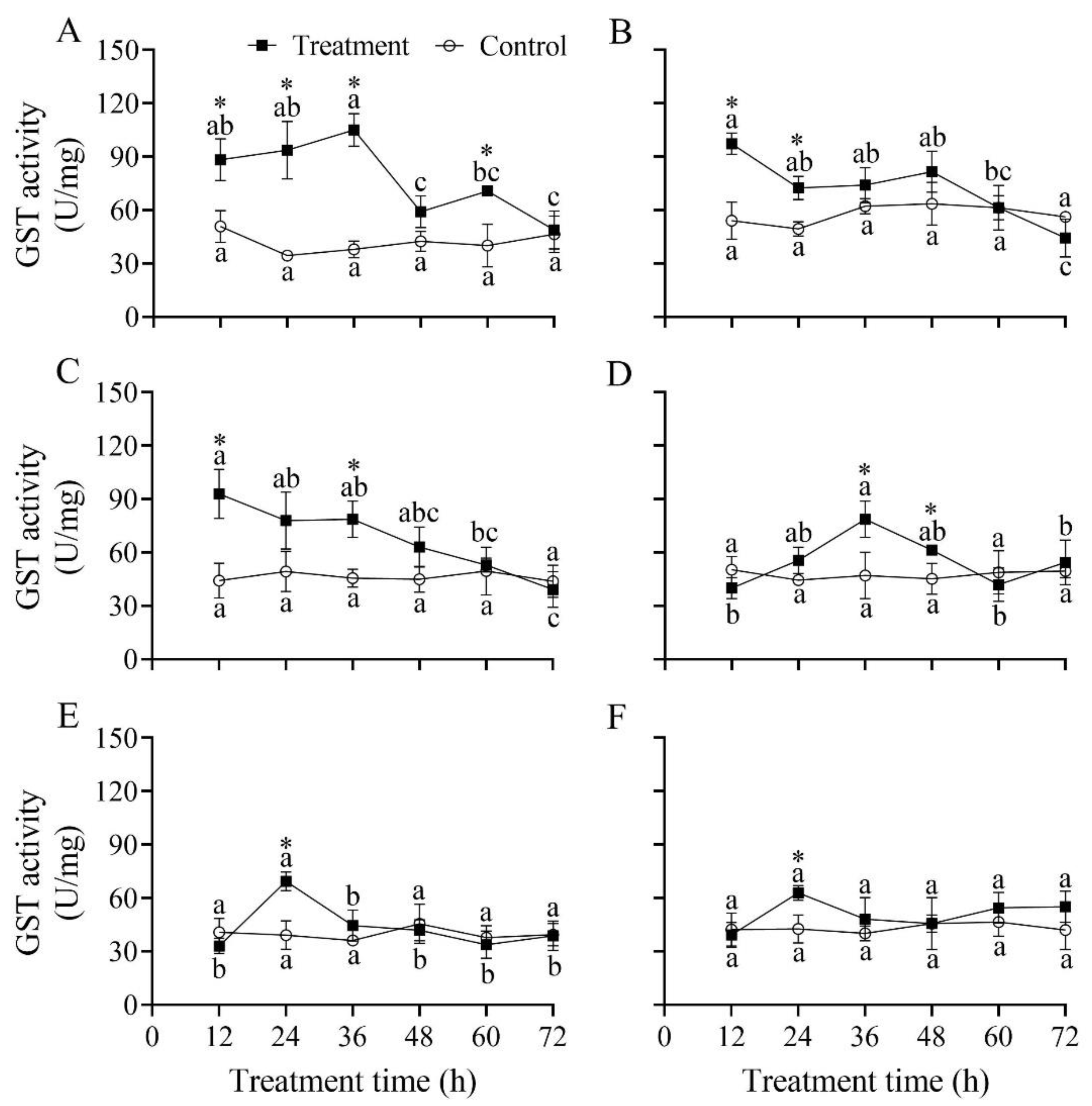
3.9. Effect of B. bassiana PfBb on GST Enzyme Activity in S. frugiperda Larvae
3.10. Effect of B. bassiana PfBb on CarE Enzyme Activity in S. frugiperda Larvae
3.11. Effect of B. bassiana PfBb on CYP450 Enzyme Activity in S. frugiperda Larvae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, X.N.; Jin, D.C.; Zou, X.; Guo, J.J. Laboratory and field evaluation of an entomopathogenic fungus, Isaria cateniannulata strain 08XS-1, against Tetranychus urticae (Koch). Pest Manag. Sci. 2016, 72, 1059–1066. [Google Scholar] [CrossRef] [Green Version]
- Ghaffari, S.; Karimi, J.; Kamali, S. Moghadam EM. Biocontrol of Planococcus citri (Hemiptera: Pseudococcidae) by Lecanicillium longisporum and Lecanicillium lecanii under laboratory and greenhouse conditions. J. Asia-Pac. Entomol. 2017, 20, 605–612. [Google Scholar] [CrossRef]
- Wasuwan, R.; Phosrithong, N.; Promdonkoy, B.; Sangsrakru, D.; Sonthirod, C.; Tangphatsornruang, S.; Likhitrattanapisal, S.; Ingsriswang, S.; Srisuksam, C.; Klamchao, K.; et al. The fungus Metarhizium sp. BCC 4849 is an effective and safe mycoinsecticide for the management of spider mites and other insect pests. Insects 2021, 13, 42. [Google Scholar] [CrossRef] [PubMed]
- Zemek, R.; Konopická, J.; Jozová, E.; Skoková, H.O. Virulence of Beauveria bassiana strains isolated from cadavers of Colorado potato beetle, Leptinotarsa decemlineata. Insects 2021, 12, 1077. [Google Scholar] [CrossRef] [PubMed]
- Clifton, E.H.; Hajek, A.E.; Jenkins, N.E.; Roush, R.T.; Rost, J.P.; Biddinger, D.J. Applications of Beauveria bassiana (Hypocreales: Cordycipitaceae) to control populations of spotted lanternfly (Hemiptera: Fulgoridae), in semi-natural landscapes and on grapevines. Environ. Entomol. 2020, 49, 854–864. [Google Scholar] [CrossRef]
- Wang, H.H.; Liu, S.; Wang, S.Y.; Lei, Z.R. Research and development of wettable powder of Beauveria bassiana and its control and application to Frankliniella occidentalis. Chin. J. Biol. Control 2020, 36, 858–861. (In Chinese) [Google Scholar] [CrossRef]
- Sullivan, C.F.; Parker, B.L.; Skinner, M. A Review of commercial Metarhizium- and Beauveria-based biopesticides for the biological control of ticks in the USA. Insects 2022, 13, 260. [Google Scholar] [CrossRef]
- Ma, X.M.; Liu, X.X.; Ning, X.; Zhang, B.; Han, F.; Guan, X.M.; Tan, Y.F.; Zhang, Q.W. Effects of Bacillus thuringiensis toxin Cry1Ac and Beauveria bassiana on Asiatic corn borer (Lepidoptera: Crambidae). J. Invertebr. Pathol. 2008, 99, 123–128. [Google Scholar] [CrossRef]
- Reddy, G.V.P.; Zhao, Z.H.; Humber, R.A. Laboratory and field efficacy of entomopathogenic fungi for the management of the sweetpotato weevil, Cylas formicarius (Coleoptera: Brentidae). J. Invertebr. Pathol. 2014, 122, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Zafar, J.; Freed, S.; Khan, B.A.; Farooq, M. Effectiveness of Beauveria bassiana against cotton whitefly, Bemisia tabaci (Gennadius) (Aleyrodidae: Homoptera) on different host plants. Pak. J. Zool. 2016, 48, 91–99. Available online: http://zsp.com.pk/pdf48/91-99%20(11)%20PJZ-2260-15%2011-11-15%20Effectiveness%20of%20Beauveria%20bassiana%20against%20cotton%20wh_.pdf (accessed on 5 April 2022).
- Fargues, J.; Remaudiere, G. Considerations on the specificity of entomopathogenic fungi. Mycopathologia 1977, 62, 31–37. Available online: https://link.springer.com/article/10.1007/BF00491993 (accessed on 26 June 2022). [CrossRef]
- Cao, W.P.; Song, J.; Zhen, W.; Wang, J.Y.; Feng, S.L.; Du, L.X. Correlation between biological characteristics of Beauveria bassiana and its cuticle infection on different insects. Chin. J. Biol. Control 2013, 29, 503–508. (In Chinese) [Google Scholar] [CrossRef]
- Liu, J.; Ling, Z.Q.; Wang, J.J.; Xiang, T.T.; Xu, L.; Gu, C.X.; Liu, R.; Xu, J.; Xu, C.L.; Zhou, W.; et al. In vitro transcriptomes analysis identifies some special genes involved in pathogenicity difference of the Beauveria bassiana against different insect hosts. Microb. Pathogenesis. 2021, 154, 104824. [Google Scholar] [CrossRef]
- Mazet, I.; Boucias, D.G. Effects of the fungal pathogen, Beauveria bassiana on protein biosynthesis of infected Spodoptera exigua larvae. J. Insect Physiol. 1996, 42, 91–99. [Google Scholar] [CrossRef]
- Qin, G.H.; Jia, M.; Liu, T.; Xuan, T.; Zhu, K.Y.; Guo, Y.P.; Ma, E.B.; Zhang, J.Z. Identification and characterisation of ten glutathione S-transferase genes from oriental migratory locust, Locusta migratoria manilensis (Meyen). Pest Manag. Sci. 2011, 67, 697–704. [Google Scholar] [CrossRef]
- Newcomb, R.D.; Campbell, P.M.; Ollis, D.L.; Cheah, E.; Russell, R.J.; Oakeshott, J.G. A single amino acid substitution converts a carboxylesterase to an organophosphorus hydrolase and confers insecticide resistance on a blowfly. Proc. Natl. Acad. Sci. USA 1997, 94, 7464–7468. [Google Scholar] [CrossRef] [Green Version]
- Cheng, T.C.; Wu, J.Q.; Wu, Y.Q.; Chilukuri, R.V.; Huang, L.H.; Yamamoto, K.; Feng, L.; Li, W.S.; Chen, Z.W.; Guo, H.Z.; et al. Genomic adaptation to polyphagy and insecticides in a major East Asian noctuid pest. Nat. Ecol. Evol. 2017, 1, 1747–1756. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, J.P.; Bailey, A.M.; Cobb, B.; Vilcinskas, A. Fungi as elicitors of insect immune responses. Arch. Insect Biochem. Physiol. 2000, 44, 49–68. [Google Scholar] [CrossRef]
- Ding, J.N.; Zhang, H.H.; Chi, D.F. Effects of a pathogenic Beauveria bassiana (Hypocreales: Cordycipitaceae) strain on detoxifying and protective enzyme activities in Xylotrechus rusticus (Coleoptera: Cerambycidae) larvae. Fla. Entomol. 2015, 98, 1148–1156. [Google Scholar] [CrossRef] [Green Version]
- Dubovskiy, I.M.; Martemyanov, V.V.; Vorontsova, Y.L.; Rantala, M.J.; Gryzanova, E.V.; Glupov, V.V. Effect of bacterial infection on antioxidant activity and lipid peroxidation in the midgut of Galleria mellonella L. larvae (Lepidoptera, Pyralidae). Comp. Biochem. Phys. C 2008, 148, 1–5. [Google Scholar] [CrossRef]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Ki, Y.W.; Lee, J.E.; Park, J.H.; Shin, I.C.; Koh, H.C. Reactive oxygen species and mitogen-activated protein kinase induce apoptotic death of SH-SY5Y cells in response to fipronil. Toxicol. Lett. 2012, 211, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Li, P.W.; Fu, X.L.; Chen, S.C.; Hu, X.; Wang, X.Q.; Peng, P. Effect of temperature stress on four protective enzymes and overall antioxidant capacity in Darna trima (Moore). Chin. J. Appl. Entomol. 2016, 53, 809–816. (In Chinese) [Google Scholar] [CrossRef]
- Jayanthi, P.D.K.; Arthikirubha, A.; Vivek, K.; Ravindra, M.A.; Selvakumar, G.; Abraham, V. Aspergillus flavus impairs antioxidative enzymes of Sternochetus mangiferae during mycosis. J. Invertebr. Pathol. 2015, 124, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Li, G.N.; Shi, M.; Zhao, S.; Long, Y.H.; Zhu, Y. Toxicity response of silkworm intestine to Bacillus cereus SW7-1 pathogen. Sci. Total Environ. 2019, 692, 1282–1290. [Google Scholar] [CrossRef] [PubMed]
- Sparks, A.N. A review of the biology of the fall armyworm. Fla. Entomol. 1979, 62, 82–87. [Google Scholar] [CrossRef]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamo, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (J.E. Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in west and central africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [Green Version]
- Sharanabasappa; Kalleshwaraswamy, C.M.; Asokan, R.; Mahadeva, S.H.M.; Maruthi, M.S.; Pavithra, H.B.; Hegde, K.; Navi, S.; Prabhu, S.T.; Goergen, G. First report of the fall armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae), an alien invasive pest on maize in India. Pest Manag. Horticul. Ecosyst. 2018, 24, 23–29. Available online: http://aapmhe.in/index.php/pmhe/article/view/816 (accessed on 4 June 2022).
- Guo, J.F.; Zhao, J.Z.; He, K.L.; Zhang, F.; Wang, Z.Y. Potential invasion of the crop-devastating insect pest fall armyworm Spodoptera frugiperda to China. Plant Protection. 2018, 44, 1–10. (In Chinese) [Google Scholar] [CrossRef]
- Chen, H.; Yang, X.L.; Chen, A.D.; Li, Y.C.; Wang, D.H.; Liu, J.; Hu, G. Immigration timing and origin of the first fall armyworms (Spodoptera frugiperda) detected in China. Chin. J. Appl. Entomol. 2020, 57, 1270–1278. (In Chinese) [Google Scholar] [CrossRef]
- Wang, L.; Chen, K.W.; Zhong, G.H.; Xian, J.D.; He, X.F.; Lu, Y.Y. Progress for occurrence and management and the strategy of the fall armyworm Spodoptera frugiperda (Smith). J. Environ. Entomol. 2019, 41, 479–487. (In Chinese) [Google Scholar] [CrossRef]
- Chen, W.B.; Li, Y.Y.; Wang, M.Q.; Liu, C.X.; Mao, J.J.; Chen, H.Y.; Zhang, L.S. Natural enemy insect resources of the fall armyworm Spodoptera frugiperda, their application status, and existing problems and suggestions. Chin. J. Biol. Control 2019, 35, 658–673. (In Chinese) [Google Scholar] [CrossRef]
- Burtet, L.M.; Bernardi, O.; Melo, A.A.; Pes, M.P.; Strahl, T.T.; Guedes, J.V.C. Managing fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), with Bt maize and insecticides in southern Brazil. Pest Manag. Sci. 2017, 73, 2569–2577. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.J.; Yuan, H.Z.; Chen, Y.X.; Shi, X.; Liu, X.H.; Wang, Z.Y.; Liu, Y.; Yang, D.B. Broadcasting of tiny granules by drone to mimic liquid spraying for the control of fall armyworm (Spodoptera frugiperda). Pest Manag. Sci. 2022, 78, 43–51. [Google Scholar] [CrossRef]
- Xu, Y.D.; Wei, H.S.; Shi, J.W.; Chen, H.H.; Shi, W.P.; Tan, S.Q. Comparison of virulence of three Beauveria bassiana strains against fall armyworm Spodoptera frugiperda. J. Plant Prot. 2020, 47, 867–874. (In Chinese) [Google Scholar] [CrossRef]
- Rajula, J.; Pittarate, S.; Suwannarach, N.; Kumla, J.; Ptaszynska, A.A.; Thungrabeab, M.; Mekchay, S.; Krutmuang, P. Evaluation of native entomopathogenic fungi for the control of fall armyworm (Spodoptera frugiperda) in Thailand: A sustainable way for eco-friendly agriculture. J. Fungi 2021, 7, 1073. [Google Scholar] [CrossRef]
- Deng, J.D.; Zhuang, W.X.; Liu, Y.J.; Song, L.W.; Zhang, L.W. Pathogenicity of white muscardine fungus Beauveria bassiana against Japanese pine sawyer beetle Monochamus alternatus and its compatibility with ectoparasitic beetle Dastarcus helophoroides. J. Plant Prot. 2021, 48, 602–609. (In Chinese) [Google Scholar] [CrossRef]
- Akutse, K.S.; Kimemia, J.W.; Ekesi, S.; Khamis, F.M.; Ombura, O.L.; Subramanian, S. Ovicidal effects of entomopathogenic fungal isolates on the invasive fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Appl. Entomol. 2019, 143, 626–634. [Google Scholar] [CrossRef]
- Faria, M.; Souza, D.A.; Sanches, M.M.; Schmidt, F.G.V.; Oliveira, C.M.; Benito, N.P.; Lopes, R.B. Evaluation of key parameters for developing a Metarhizium rileyi-based biopesticide against Spodoptera frugiperda (Lepidoptera: Noctuidae) in maize: Laboratory, greenhouse, and field trials. Pest Manag. Sci. 2021, 78, 1146–1154. [Google Scholar] [CrossRef]
- Idrees, A.; Qadir, Z.A.; Akutse, K.S.; Afzal, A.; Hussain, M.; Islam, W.; Waqas, M.S.; Bamisile, B.S.; Li, J. Effectiveness of entomopathogenic fungi on immature stages and feeding performance of fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) Larvae. Insects 2021, 12, 1044. [Google Scholar] [CrossRef]
- Chen, X.M.; Wang, X.Y.; Lu, W.; Zheng, X.L. Virulence of four entomogenous fungi against the larvae of Phauda flammans (Walker). Guangxi Plant Prot. 2020, 33, 1–5. Available online: https://t.cnki.net/kcms/detail?v=2Hzf_72mFvAinGMY72vEySGSVRX8f3T-EUYTkh75dxlmRCNh_FcTRWY2UnAX-wTlzeaPgnO2fsU0Y4R245ZN3YNAV9_3U3kExfojCOcmXzFNw6Gm1p0xZWTbaJfcJkGK&uniplatform=NZKPT (accessed on 25 May 2021). (In Chinese).
- Chen, X.M.; Wang, X.Y.; Lu, W.; Zheng, X.L. Use of Beauveria bassiana in combination with commercial insecticides to manage Phauda flammans (Walker) (Lepidoptera: Phaudidae): Testing for compatibility and synergy. J. Asia-Pac. Entomol. 2021, 24, 272–278. [Google Scholar] [CrossRef]
- Qasim, M.; Lin, Y.W.; Dash, C.K.; Bamisile, B.S.; Ravindran, K.; Ul Islam, S.; Ali, H.; Wang, F.F.; Wang, L.D. Temperature-dependent development of Asian citrus psyllid on various hosts, and mortality by two strains of Isaria. Microb. Pathogenesis 2018, 119, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.G.; Liu, C.L.; Xu, J.H.; Xu, Q. Modeling and biological implication of time-dose-mortality data for the entomophthoralean fungus, Zoophthora anhuiensis, on the green peach aphid Myzus persicae. J. Invertebr Pathol. 1998, 72, 246–251. [Google Scholar] [CrossRef]
- Qasim, M.; Xiao, H.M.; He, K.; Omar, M.A.A.; Hussain, D.; Noman, A.; Rizwan, M.; Khan, K.A.; Al-zoubi, O.M.; Alharbi, S.A.; et al. Host-pathogen interaction between Asian citrus psyllid and entomopathogenic fungus (Cordyceps fumosorosea) is regulated by modulations in gene expression, enzymatic activity and HLB-bacterial population of the host. Comp. Biochem. Phys. C 2021, 248, 109112. [Google Scholar] [CrossRef] [PubMed]
- El Husseini, M.M.M. Effect of the fungus, Beauveria bassiana (Balsamo) Vuillemin, on the beet armyworm, Spodoptera exigua (Hübner) larvae (Lepidoptera: Noctuidae), under laboratory and open field conditions. Egypt. J. Biol. Pest Control 2019, 29, 52. [Google Scholar] [CrossRef] [Green Version]
- Omar, G.; Ibrahim, A.; Hamadah, K. Virulence of Beauveria bassiana and Metarhizium anisopliae on different stages of the pink bollworm, Pectinophora gossypiella (Saunders) (Lepidoptera: Gelechiidae). Egypt. J. Biol. Pest Control 2021, 31, 102. [Google Scholar] [CrossRef]
- Schulte, M.J.; Martin, K.; Büchse, A.; Sauerborn, J. Entomopathogens (Beauveria bassiana and Steinernema carpocapsae) for biological control of bark-feeding moth Indarbela dea on field-infested litchi trees. Pest Manag. Sci. 2009, 65, 105–112. [Google Scholar] [CrossRef]
- Ji, X.Y.; Yang, C.J. Infection and application of Beauveria spp. Chin. J. Biol. Control 2003, 19, 82–85. (In Chinese) [Google Scholar] [CrossRef]
- Butt, T.M.; Ibrahim, L.; Clark, S.J.; Beckett, A. The germination behaviour of Metarhizium anisopliae on the surface of aphid and flea beetle cuticles. Mycol. Res. 1995, 99, 945–950. [Google Scholar] [CrossRef]
- Wilson, K.; Cotter, S.C.; Reeson, A.F.; Pell, J.K. Melanism and disease resistance in insects. Ecol. Lett. 2011, 4, 637–649. [Google Scholar] [CrossRef] [Green Version]
- Mannino, M.C.; Huarte-Bonnet, C.; Davyt-Colo, B.; Pedrini, N. Is the insect cuticle the only entry gate for fungal infection? insights into alternative modes of action of entomopathogenic fungi. J. Fungi 2019, 5, 33. [Google Scholar] [CrossRef] [Green Version]
- Lü, L.H.; He, Y.R.; Wu, Y.J.; Feng, X.; Chen, H.Y. The time-dose-mortality model of a Paecilomyces fumosoroseus isolate on the diamondback moth, Plutella xylostella. Acta Entomol. Sin. 2007, 50, 567–573. (In Chinese) [Google Scholar] [CrossRef]
- He, L.M.; Ge, S.S.; Chen, Y.C.; Wu, Q.L.; Jiang, Y.Y.; Wu, K.M. The developmental threshold temperature, effective accumulated temperature and prediction model of developmental duration of fall armyworm, Spodoptera frugiperda. Plant Prot. 2019, 45, 18–26. (In Chinese) [Google Scholar] [CrossRef]
- Jiang, X.C.; Shen, Y.D.; Sun, J.C.; Li, X.X.; Huang, Y.; Dong, Y.C.; Cao, H.Q. Effect of chlorantraniliprole and emamectin benzoate on toxicity and detoxification enzymes activity in Spodoptera frugiperda larva. J. Environ. Entomol. 2019, 41, 961–967. (In Chinese) [Google Scholar] [CrossRef]
- Wen, S.F.; Xue, Y.N.; Du, R.S.; Liu, C.; Wang, X.T.; Wang, Y.W.; Liu, C.; Wang, S.; Wang, J.H.; Xia, X.M. Toxicity and sublethal effects of triflumezopyrim on the development and detoxification enzymatic activities in the small brown planthopper (SBPH), Laodelphax striatellus (Fallen). Crop Prot. 2021, 150, 105813. [Google Scholar] [CrossRef]
- Wang, H.M.; Zhang, H.; Hao, C.; Zhang, X.H. Effects of Isaria fumosorosea infection on different enzyme activities in the larvae of Plutella xylostella. Mycosystema 2013, 32, 269–276. (In Chinese) [Google Scholar] [CrossRef]
- Li, Z.Z.; Shen, H.J.; Jiang, Q.G.; Ji, B.Z. A study on the activity of endogenous enzymes of protective system in some insects. Acta Entomol. Sin. 1994, 37, 399–403. (In Chinese) [Google Scholar] [CrossRef]
- Tian, J.; Diao, H.L.; Liang, L.; Hao, C.; Arthurs, S.; Ma, R.Y. Pathogenicity of Isaria fumosorosea to Bemisia tabaci, with some observations on the fungal infection process and host immune response. J. Invertebr Pathol. 2015, 130, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chen, S.B.; Wu, C.Y.; Zhang, B.Y.; Zhang, Y.; Teng, B.; Hu, B.J. Screening of Beauveria bassiana strains with high virulence against armyworm and activities of protective enzymes in the larvae infected by fungi. J. Nucl. Agr. Sci. 2020, 34, 2701–2707. Available online: https://www.hnxb.org.cn/CN/10.11869/j.issn.100-8551.2020.12.2701 (accessed on 8 March 2022). (In Chinese).
- Wang, S.C.; He, R.F.; Lu, M.Q.; Huang, H.P.; Wang, N.Y.; Guo, X.J.; Geng, T. Effects on antioxidant levels of silkworm larvae infected with Beauveria bassiana. Chin. J. Trop. Crops. 2017, 38, 2136–2144. (In Chinese) [Google Scholar] [CrossRef]
- Felton, G.W.; Summers, C.B. Antioxidant systems in insects. Arch. Insect Biochem. 1995, 29, 187–197. [Google Scholar] [CrossRef]
- Wang, Y.; Oberley, L.W.; Murhammer, D.W. Antioxidant defense systems of two lipidopteran insect cell lines. Free Radic. Biol. Med. 2001, 30, 1254–1262. [Google Scholar] [CrossRef]
- Wu, Q.J.; Zhang, Y.J.; Xu, B.Y.; Zhang, W.J. The defending enzymes in abamectin resistant Plutella xylostella. Chin. J. Appl. Entomol. 2011, 48, 291–295. Available online: http://www.ent-bull.com.cn/downfile.aspx?qi_id=609&id=15159 (accessed on 15 May 2022). (In Chinese).
- Shi, G.L.; Wang, Y.N.; Wang, H.L.; Zhao, L.L.; Liu, S.Q.; Cao, H.; Yu, T.Q.; Lu, P. Effects of Tagetes erecta extracts on glutathione S transferase and protease activities and protein content in Tetranychus viennensis. Chin. J. Appl. Ecol. 2007, 18, 400–404. Available online: http://www.cjae.net/CN/Y2007/V18/I02/400 (accessed on 27 April 2022). (In Chinese).
- Chen, X.M.; Wang, X.Y.; Lu, W.; Zheng, X.L. Effects of Beauveria bassiana on the larval protein content, protective and detoxification enzyme activities in Phauda flammans (Lepidoptera: Phaudidae). J. Plant Prot. 2020, 47, 1379–1380. (In Chinese) [Google Scholar] [CrossRef]
- Chavez, V.M.; Marques, G.; Delbecque, J.P.; Kobayashi, K.; Hollingsworth, M.; Burr, J.; Natzle, J.E.; O’Connor, M.B. The Drosophila disembodied gene controls late embryonic morphogenesis and codes for a cytochrome P450 enzyme that regulates embryonic ecdysone levels. Development 2000, 127, 4115–4126. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, J.C.; Lee, S.J.; Lee, M.R.; Park, S.E.; Li, D.; Baek, S.; Shin, T.Y.; Kim, J.S. Beauveria bassiana ERL836 and JEF-007 with similar virulence show different gene expression when interacting with cuticles of western flower thrips, Frankniella occidentalis. BMC Genom. 2020, 21, 836. [Google Scholar] [CrossRef] [PubMed]
- Ismail, H.M.; Freed, S.; Naeem, A.; Malik, S.; Ali, N. The effect of entomopathogenic fungi on enzymatic activity in chlorpyrifos-resistant mosquitoes, Culex quinquefasciatus (Diptera: Culicidae). J. Med. Entomol. 2020, 57, 204–213. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Instar | Regression Equation | χ2 | p | LC50 (×105 Spores/mL) | 95% Confidence Limit (Spores/mL) |
|---|---|---|---|---|---|
| First | y = 0.42x − 2.48 | 0.46 | 0.796 | 7.68 | 2.13 × 105~1.92 × 106 |
| Second | y = 0.54x − 3.66 | 1.23 | 0.541 | 54.64 | 2.62 × 106~1.23 × 107 |
| Third | y = 0.44x − 3.22 | 0.36 | 0.836 | 220.11 | 8.75 × 106~8.88 × 107 |
| Fourth | y = 0.40x − 3.43 | 0.50 | 0.781 | 3084.06 | 7.08 × 107~7.69 × 109 |
| Fifth | y = 0.41x − 3.67 | 0.78 | 0.678 | 9579.65 | 1.62 × 108~6.91 × 1010 |
| Sixth | y = 0.23x − 2.62 | 0.50 | 0.780 | 2548351.52 | 1.66 × 109~1.49 × 1031 |
| Instar | Conditional Mortality Model | Cumulative Mortality Model | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Parameter | Value | SE | t | P | Parameter | Value | Var (τ) | Cov (β, τ) | |
| First | β | 0.41 | 0.14 | 2.99 | 0.010 | β | 0.41 | 0.00 | 0.00 |
| γ1 | −4.35 | 0.98 | 4.46 | 0.001 | τ1 | −4.35 | 0.06 | −0.01 | |
| γ2 | −4.21 | 0.97 | 4.32 | 0.001 | τ2 | −3.59 | 0.06 | −0.01 | |
| γ3 | −4.11 | 0.97 | 4.23 | 0.001 | τ3 | −3.12 | 0.06 | −0.01 | |
| γ4 | −4.24 | 0.99 | 4.28 | 0.001 | τ4 | −2.84 | 0.06 | −0.01 | |
| γ5 | −5.55 | 1.18 | 4.70 | 0.000 | τ5 | −2.77 | 0.06 | −0.01 | |
| Second | β | 0.58 | 0.17 | 3.53 | 0.003 | β | 0.58 | 0.00 | 0.00 |
| γ1 | −5.82 | 1.20 | 4.83 | 0.000 | τ1 | −5.82 | 0.25 | −0.03 | |
| γ2 | −5.88 | 1.22 | 4.82 | 0.000 | τ2 | −5.15 | 0.24 | −0.03 | |
| γ3 | −5.59 | 1.19 | 4.69 | 0.000 | τ3 | −4.65 | 0.24 | −0.03 | |
| γ4 | −6.18 | 1.26 | 4.91 | 0.000 | τ4 | −4.46 | 0.23 | −0.03 | |
| γ5 | −6.84 | 1.34 | 5.10 | 0.000 | τ5 | −4.37 | 0.23 | −0.03 | |
| Third | β | 0.56 | 0.18 | 3.07 | 0.007 | β | 0.56 | 0.00 | 0.00 |
| γ1 | −6.25 | 1.38 | 4.54 | 0.000 | τ1 | −6.25 | 0.27 | −0.03 | |
| γ2 | −5.89 | 1.32 | 4.47 | 0.000 | τ2 | −5.36 | 0.25 | −0.03 | |
| γ3 | −6.03 | 1.35 | 4.47 | 0.000 | τ3 | −4.95 | 0.24 | −0.03 | |
| γ4 | −6.19 | 1.36 | 4.56 | 0.000 | τ4 | −4.69 | 0.24 | −0.03 | |
| γ5 | −6.53 | 1.38 | 4.74 | 0.000 | τ5 | −4.55 | 0.24 | −0.03 | |
| γ6 | −8.00 | 1.83 | 4.38 | 0.000 | τ6 | −4.52 | 0.24 | −0.03 | |
| Fourth | β | 0.61 | 0.25 | 2.50 | 0.023 | β | 0.61 | 0.01 | 0.01 |
| γ1 | −7.36 | 1.90 | 3.88 | 0.001 | τ1 | −7.36 | 0.55 | −0.07 | |
| γ2 | −7.29 | 1.86 | 3.92 | 0.001 | τ2 | −6.63 | 0.51 | −0.07 | |
| γ3 | −6.95 | 1.81 | 3.85 | 0.001 | τ3 | −6.09 | 0.49 | −0.07 | |
| γ4 | −6.79 | 1.79 | 3.80 | 0.001 | τ4 | −5.68 | 0.48 | −0.07 | |
| γ5 | −7.28 | 1.80 | 4.05 | 0.001 | τ5 | −5.50 | 0.47 | −0.07 | |
| γ6 | −9.34 | 2.61 | 3.58 | 0.002 | τ6 | −5.48 | 0.47 | −0.07 | |
| Fifth | β | 0.70 | 0.29 | 2.39 | 0.029 | β | 0.70 | 0.02 | 0.02 |
| γ1 | −8.65 | 2.35 | 3.68 | 0.002 | τ1 | −8.65 | 1.32 | −0.15 | |
| γ2 | −8.01 | 2.22 | 3.61 | 0.002 | τ2 | −7.58 | 1.17 | −0.15 | |
| γ3 | −8.36 | 2.24 | 3.73 | 0.002 | τ3 | −7.21 | 1.14 | −0.15 | |
| γ4 | −7.37 | 2.12 | 3.49 | 0.003 | τ4 | −6.59 | 1.09 | −0.15 | |
| γ5 | −8.64 | 2.21 | 3.91 | 0.001 | τ5 | −6.47 | 1.07 | −0.15 | |
| γ6 | −8.60 | 2.23 | 3.86 | 0.001 | τ6 | −6.36 | 1.07 | −0.15 | |
| Sixth | β | 0.47 | 0.33 | 1.42 | 0.173 | β | 0.47 | 0.03 | 0.03 |
| γ2 | −6.70 | 2.52 | 2.66 | 0.016 | τ2 | −6.70 | 1.83 | −0.23 | |
| γ3 | −7.23 | 2.58 | 2.80 | 0.012 | τ3 | −6.23 | 1.78 | −0.23 | |
| γ4 | −6.79 | 2.40 | 2.83 | 0.012 | τ4 | −5.78 | 1.68 | −0.22 | |
| γ5 | −6.34 | 2.37 | 2.67 | 0.016 | τ5 | −5.33 | 1.63 | −0.22 | |
| γ6 | −7.12 | 2.35 | 3.03 | 0.008 | τ6 | −5.18 | 1.59 | −0.22 | |
| γ7 | −8.51 | 2.90 | 2.94 | 0.009 | τ7 | −5.14 | 1.58 | −0.22 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.-P.; Luo, M.; Wang, X.-Y.; He, X.Z.; Lu, W.; Zheng, X.-L. Pathogenicity of Beauveria bassiana PfBb and Immune Responses of a Non-Target Host, Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2022, 13, 914. https://doi.org/10.3390/insects13100914
Gao Y-P, Luo M, Wang X-Y, He XZ, Lu W, Zheng X-L. Pathogenicity of Beauveria bassiana PfBb and Immune Responses of a Non-Target Host, Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects. 2022; 13(10):914. https://doi.org/10.3390/insects13100914
Chicago/Turabian StyleGao, Yi-Ping, Mei Luo, Xiao-Yun Wang, Xiong Zhao He, Wen Lu, and Xia-Lin Zheng. 2022. "Pathogenicity of Beauveria bassiana PfBb and Immune Responses of a Non-Target Host, Spodoptera frugiperda (Lepidoptera: Noctuidae)" Insects 13, no. 10: 914. https://doi.org/10.3390/insects13100914