
Local Competition and Enhanced Defense: How Metarhizium brunneum Inhibits Verticillium longisporum in Oilseed Rape Plants


Agricultural Entomology, Department of Crop Sciences, University of Goettingen, Grisebachstr 6, 37077 Goettingen, Germany
*
Authors to whom correspondence should be addressed.
J. Fungi 2023, 9(8), 796; https://doi.org/10.3390/jof9080796
Submission received: 30 May 2023
/
Revised: 27 June 2023
/
Accepted: 21 July 2023
/
Published: 28 July 2023
(This article belongs to the Special Issue Plant and Fungal Interactions, 2nd Edition)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Metarhizium brunneum is a soil-borne fungal entomopathogen that can be associated with plant roots. Previous studies have demonstrated that root colonization by beneficial fungi can directly affect soil-borne pathogens through competition and antibiosis and can activate a systemic response in plants, resulting in a primed state for a faster and/or stronger response to stressors. However, the mechanisms by which Metarhizium inoculation ameliorates symptoms caused by plant pathogens are not well known. This study evaluated the ability of M. brunneum to protect oilseed rape (Brassica napus L.) plants against the soil-borne pathogen Verticillium longisporum and investigated whether the observed effects are a result of direct interaction and/or plant-mediated effects. In vitro and greenhouse experiments were conducted to measure fungal colonization of the rhizosphere and plant tissues, and targeted gene expression analysis was used to evaluate the plant response. The results show that M. brunneum delayed pathogen colonization of plant root tissues, resulting in decreased disease symptoms. Direct competition and antibiosis were found to be part of the mechanisms, as M. brunneum growth was stimulated by the pathogen and inhibited the in vitro growth of V. longisporum. Additionally, M. brunneum changed the plant response to the pathogen by locally activating key defense hormones in the salicylic acid (SA) and abscisic acid (ABA) pathways. Using a split-root setup, it was demonstrated that there is a plant-mediated effect, as improved plant growth and decreased disease symptoms were observed when M. brunneum was in the systemic compartment. Moreover, a stronger systemic induction of the gene PR1 suggested a priming effect, involving the SA pathway. Overall, this study sheds light on the mechanisms underlying the protective effects of M. brunneum against soil-borne pathogens in oilseed rape plants, highlighting the potential of this fungal entomopathogen as a biocontrol agent in sustainable agriculture.
1. Introduction
Roots grow in the soil and share the same substrate with numerous microbes. It has been estimated that a gram of soil can contain up to 10 billion microorganisms [1]. Moreover, plant roots deposit up to 40% of the carbon they fix during photosynthesis into the soil. This process, called rhizodeposition, creates a microenvironment that promotes a 10- to 100-fold increase in microbial density around plant roots and a microbial community composition that is distinct from that found in bulk soil [2,3]. This microenvironment, known as rhizosphere, is the site for multiple interactions between soil microbes and plant roots. Soil microorganisms can have beneficial effects on plants as they can facilitate nutrient uptake or protect against abiotic and biotic stresses [4,5].
Several Metarhizium species, primarily known as fungal pathogens of insects, are also rhizosphere competent. They exhibit enhanced growth around roots [6,7] and can colonize plant tissues [7,8,9]. Together with other genera of entomopathogens such as Beauveria and Lecanicillium, these fungi have been referred to as “endophytic entomopathogenic fungi” (EEF). Several studies have reported positive effects of EEF on plants, including ameliorating salt stress [10,11], promoting growth [12,13,14], improving nutrient acquisition [15,16,17] and reducing insect and pathogen damage [18,19].
In the past decade, an increasing number of studies have demonstrated that different Metarhizium species can inhibit several plant pathogens in vitro [20,21,22]. Moreover, colonization by Metarhizium can reduce pathogen-induced symptoms in various plants [20,21,22,23,24]. However, the mechanisms by which Metarhizium species antagonize plant pathogens are not well understood [25,26]. Plant protection can result from a combination of competition and antibiosis [20,21,22]. Metarhizium has been shown to strongly inhibit the growth of pathogens in vitro through the production of secondary metabolites that are toxic to microorganisms and insects [27]. Mycoparasitism has been suggested as another potential mechanism, although this has not been observed with Metarhizium [28]. Another mechanism suggested is the activation of plant defenses. Beneficial microorganisms can trigger induced systemic resistance (ISR) in plants [4], which activates plant defenses and helps protect against pathogens. This has been observed with obligate symbionts such as mycorrhiza [29,30] and facultative endophytes such as Trichoderma or other non-pathogenic fungal strains [4,31].
Some studies have shown that EEF may modify plant defense pathways. For instance, peanut plants inoculated with M. anisopliae strain M202-1 showed downregulation of genes involved in the hypersensitive response and synthesis of resistance proteins [32]. Increased defense responses have also been observed. The jasmonic acid (JA) and salicylic acid (SA) pathways are essential in the defense response and were activated by B. bassiana BG11 in Arabidopsis thaliana [33] and by the strains Bb0062 and Bb02 in Nicotiana benthamiana [34]. The inoculation of M. brunneum Mb7 induced the expression of pathogen resistance genes such as PR1 that play a role in the SA pathway [24]. Levels of SA and JA hormones were also higher in oilseed rape plants inoculated with M. brunneum F52 [35] and in maize plants inoculated with M. anisopliae A1080 [36] and M. robertsii strain ARSEF 14325 [12].
Recent publications suggest that Metarhizium primes plant defense responses against insects [37,38,39]. The phenomenon of priming is associated with ISR, whereby plants exhibit a faster and/or stronger defense response when encountering a stressor if they have been previously exposed to a beneficial agent. In addition, it has been demonstrated that pre-inoculation with M. brunneum Mb7 increased reactive oxygen species production in tomato plants elicited with the fungal protein ethylene-inducing xylanase, which is an inducer of plant defense responses [24], indicating that Metarhizium could prime plants against pathogens. However, to our knowledge, there is currently little knowledge on how Metarhizium modifies plant responses towards a pathogenic fungus.
Verticillium longisporum is a soil-borne pathogen that causes Verticillium stem striping in oilseed rape (Brassica napus L. spp. oleifera) [40]. Melanized microsclerotia present in the soil germinate upon induction by plant root exudates, and the resulting hyphae grow towards the root, colonizing the surface of the root hairs and penetrating the roots through rhizodermal cells [41]. Subsequently, the hyphae grow towards the central cylinder and enter the xylem vessels, spreading into aboveground organs. Upon plant tissue death, the pathogen produces microsclerotia that can remain in plant debris in the soil for more than 10 years [40].
In this study, we investigated the interactions between M. brunneum and V. longisporum during their colonization of oilseed rape plants. Our hypothesis was that, as soil and root inhabitants, both fungi could directly affect each other’s growth and plant colonization. We also hypothesized that M. brunneum, as a plant symbiont, could modify the plant’s response to V. longisporum. To study the direct interaction, we carried out in vitro dual confrontation assays and co-inoculation of oilseed rape roots with both fungi, where we measured fungal colonization of the hypocotyl and plant disease progress. In order to distinguish direct effects from plant-mediated effects, we used a split-root experiment to further explore how each fungus affected the other’s colonization of the rhizosphere, roots, and hypocotyl. We also measured the local and systemic plant responses to each fungus by analyzing the expression of marker genes involved in plant defenses and explored whether M. brunneum could modify the plant’s response to V. longisporum.
2. Materials and Methods
2.1. Study System
Metarhizium brunneum Cb15III was obtained from the in-house collection of the Division of Agricultural Entomology. The pathogen Verticillium longisporum Vl43 was provided by the Division of Plant Pathology and Crop Protection, University of Goettingen. To obtain spore suspensions, fungi were grown on potato dextrose agar (PDA) (Carl Roth GmbH, Karlsruhe, Germany) at 23 °C for 14 days. Spores were removed from hyphae by scraping the surface of the colony with a sterile glass slide and were suspended in 20 mL of 0.1% Tween 80 (Carl Roth GmbH, Karlsruhe, Germany). The suspension was filtered through a plastic gauze and adjusted using 0.1% Tween 80 to a final concentration of 1 × 107 spores mL−1 for M. brunneum and 1.5 × 106 spores mL−1 for V. longisporum. Spore viability was assessed before each experiment by plating 100 μL of a 1 × 103 spores ml−1 distributed in 10 μL drops on PDA plates and counting the germinating colonies 48 h later.
Winter oilseed rape (Brassica napus var. Falcon) (Norddeutsche Pflanzenzucht Hans-Georg Lembke KG, NPZ, Hohenlieth, Germany) susceptible to V. longisporum [42] was used in this study. For all experiments, plants were grown in a non-sterile soil mix consisting of a mixture of commercial soil (Fruhstorfer Erde Typ 25, Hawita Gruppe GmbH, Vechta, Germany), heat-treated (steamed) compost and sand (2:1:1, v:v). Plants were kept in a greenhouse with 18–25 °C, supplemented with light to obtain a 16:8 (L:D) h photoperiod.
2.2. In Vitro Confrontation Assay
Antibiosis between M. brunneum and V. longisporum was evaluated with a confrontation assay. Single colonies of each fungal species were produced through streaking a drop of spore suspension from a glycerol stock over PDA media on a 90 mm plate. The plates were then incubated at 23 °C in the dark for 48 h. Single colonies were then transferred to a 90 mm PDA Petri dish. Each fungus was grown either alone or in dual culture with a distance of 4 cm between colonies. Plates were placed completely randomized inside the incubator. The plates were scanned at 14, 17 and 21 days after starting the confrontation assay, and the scanned images were used to measure fungal colony area with the ImageJ software (Ver. 1.53f51) [43]. Growth inhibition (GI) was calculated using the area of the colonies growing alone (control) and in dual culture with the formula:
For each treatment (single culture − dual culture × 2 fungi), 5 replicates (plates) were carried out.
2.3. In Planta Co-Inoculation Assay
To evaluate the effect of M. brunneum on the disease progress of V. longisporum infecting oilseed rape plants, a greenhouse experiment was conducted. Oilseed rape seeds were surface-sterilized in 1% sodium hypochlorite for 2 min, followed by 75% ethanol for 2 min, and rinsed three times with sterile water. Seeds were then germinated in sterile silica sand and grown for 24 days. Afterwards, seedlings were uprooted, and the root tips were cut off. The roots were then immersed in fungal spore suspension for 30 min. The treatments included the following.
- Control: roots mock-inoculated with 0.1% Tween 80.
- Mb: roots inoculated with M. brunneum Cb15III.
- Vl: roots inoculated with V. longisporum VL 43.
- Mb/Vl: roots inoculated with a mix of both fungi.
After inoculation, the seedlings were transplanted into square pots (11 × 11 cm, 0.5 L). Each treatment had 25 biological replicates, and each replicate consisted of an individual pot with a plant. The plants were arranged in a completely randomized design in a greenhouse cabin. Disease assessment, plant parameters and fungal quantification in plant hypocotyl were conducted as described below.
2.4. Split-Root Assay
A split-root system was used to differentiate effects caused by direct fungal interaction from indirect effects via induced changes in plant metabolism. Split-root ready seedlings were produced as in [37]. The inoculation of fungi was sequential to give M. brunneum time to associate with the plant roots before the inoculation with V. longisporum. Seedlings were transplanted into two bound square pots (11 cm) filled with non-sterile soil mix. One of the split-root pots was inoculated by drenching the roots with 3 mL of either M. brunneum spore suspension or 0.1% Tween 80 (mock inoculation). Seven days later, V. longisporum was inoculated by pipetting into the soil 3 mL of spore suspension. The experimental setup consisted of 5 treatments, with each plant having two root compartments (C1 and C2):
- Control: C1 = mock inoculation, C2 = untreated soil.
- Mb: C1 = M. brunneum (Mb-L), C2 = mock inoculation (Mb-S).
- VL: C1 = V. longisporum (Vl-L), C2 = mock inoculation (Vl-S).
- Local: C1 = both fungi present (Mb-L/Vl-L) C2 = both fungi absent (Mb-S/Vl-S).
- Systemic: C1 = M. brunneum present, V. longisporum absent (Mb-L/Vl-S), C2 = V. longisporum present, M. brunneum absent (VL-L/Mb-S).
“L” means local inoculation, and “S” means systemic or that the inoculation was performed in the adjacent split compartment. Each treatment had 35 plants, and plants from all treatments were randomly distributed in the greenhouse cabin.
To gather samples for gene expression analysis, a second split-root experiment was conducted. The pathogen was inoculated by root dipping to ensure better exposure to fungal spores; for this, seven days after M. brunneum inoculation, the roots of the compartment to be inoculated were carefully taken out and placed inside a 2 mL Eppendorf tube filled with V. longisporum spore suspension for 20 min, after which the roots were covered again with soil. Roots in one compartment of the control treatment had the same manipulation, dipped in 0.1% Tween 80.
2.5. Disease Assessment
In all experiments, plant height and disease severity were evaluated weekly until 35 days after fungal inoculation (co-inoculation) or V. longisporum inoculation (split-root), using the scale described in [41]:
- 1 = no symptoms.
- 2 = slight symptoms on oldest leaves (yellowing, black veins).
- 3 = slight symptoms on next younger leaves.
- 4 = about 50% of leaves showing symptoms.
- 5 = more than 50% of leaves showing symptoms.
- 6 = up to 50% of leaves dead.
- 7 = more than 50% of leaves dead.
- 8 = only apical meristem still alive.
- 9 = plant dead.
Disease severity values were used to calculate the AUDCP values (area under the disease progress curve) according to the following formula [44]:
where yi is the value of the disease severity for the “i” observation, and ti is the time after inoculation when the observation “i” was taken.
AUDPC = Σ((yi + yi+1)/2) × (ti+1 − ti)
The dry weight of roots (co-inoculation), shoots (all) and leaf area (co-inoculation) were determined at 21, 28 and 35 days post-inoculation (dpi).
2.6. Quantification of Rhizospheric and Endophytic Metarhizium brunneum
In the split-root experiment, we evaluated whether M. brunneum colonized the rhizosphere and if it were affected by V. longisporum. We measured fungal colony forming units (CFUs) in the soil closely attached to the roots at 7, 21 and 35 dpi of V. longisporum. The complete root system with soil attached was placed in 50 mL Falcon tubes (Sarstedt AG & Co. KG, Nümbrecht, Germany) with 25 mL of 0.1% Tween® 80. In order to release the rhizospheric soil from the roots, tubes were vortexed for 10 s, after which the roots were taken out for DNA extraction (see below). Tubes with rhizospheric soil were placed in a shaker in horizontal position for 20 min at 250 rpm. Samples were sonicated for 30 s, briefly vortexed and left to sediment for 20 s. Then, 100 μL of the supernatant was diluted 1:10 (v:v), and 100 μL from the dilution was plated on 9 cm Petri dishes with semi-selective medium [45]. The Petri dishes were incubated at 23 °C and 65% RH for 21 d in darkness. Fungal colonies were counted in three-day intervals, starting 10 days after plating until no new ones appeared. Colonies were confirmed as M. brunneum according to their morphology. Tubes with the original soil dilution were dried in an oven (at 60 °C for 5 days), until they reached constant weight, and the dry weight was recorded.
Fungal DNA in root (split-root experiment) and in hypocotyl (first pot experiment and split-root experiment) was measured with real-time quantitative polymerase chain reaction (qPCR). Roots were washed, dried and placed in −20 °C. For each plant, a 2 cm segment of hypocotyl was sliced, surface-sterilized in 70% ethanol for 1 min and 2% sodium hypochlorite for 5 min, rinsed three times with sterile water for 30 s and placed in −20 °C. Roots and hypocotyls were lyophilized for 72 h (Martin Christ Freeze Dryers, Osterode am Harz, Germany) and milled with a mixer mill (Retsch MM 400, Haan, Germany) in a stainless steel container with a 20 mm, 32 g steel sphere for 30 s at maximum speed. DNA was extracted from 30 mg of root tissue with the cetyltrimethylammonium bromide (CTAB) buffer extraction method described previously [46], and DNA quality was verified in agarose (0.8%) gels. The CFX384™ Real-Time System with a C1000™ Thermal Cycler (BioRad, Hercules, CA, USA) was used for fungal DNA amplification and melting curve analysis. The primers used for M. brunneum detection are specific for the Metarhizium PARB clade: Ma 1763 (CCAACTCCCAACCCC TGTGAAT) and Ma 2097 (AAAACCAGCCTCGCCGAT) [47]. Specific primers used for V. longisporum were OLG 70 (CAGCGAAACGCGATATGTAG) and OLG 71 (GGCTTGTAGGGGGTTTAGA) [41]. Amplification was performed with 1:10 dilutions of the DNA extracts. The reaction mixture contained the following: 5 μL of 2× qPCRBIO SyGreen Low-ROX (PCR Biosystems, London, UK), 0.2 μL of 10 μM of each primer and 3.6 μL of 1 μL of DNA template solution, completed to a total of 10 μL of final volume reaction. qPCR running conditions started with an initial denaturation for 2 min at 95 °C, followed by 40 reaction cycles consisting of 5 s denaturation step at 95 °C, 20 s annealing step at 66 °C (for M. brunneum) or 60 °C (for V. longisporum) and 10 s extension at 72 °C. The final elongation was performed for 5 min at 72 °C. Melting curves were obtained by heating the samples to 95 °C for 60 s and cooling them to 55 °C for 60 s followed by a temperature increase from 55 °C to 95 °C by 0.5 °C per cycle with continuous fluorescence measurement. Absolute fungal DNA amount per g of plant tissue was measured by comparing threshold cycle (Ct) values against DNA standards starting with a concentration of 100 pg × μL−1 and decreasing with a 1:3 dilution factor. The threshold cycle and standard curves were generated by the Bio-Rad CFX Maestro software (Ver. 1.1). The identity of the amplicon was verified by comparing its size with gel electrophoresis. The presence of DNA in the root was evaluated in 5 replicates (first pot experiment) or in each root compartments from 8 plant replicates (split-root experiment) for each sampling date.
2.7. Gene Expression in Plant Roots
Gene expression was analyzed in root tissues obtained from the second split-root experiment. Plants were harvested 7 days after V. longisporum inoculation as previous studies have found clear plant responses to the pathogen [48,49]. Whole roots were washed and snap-frozen in liquid nitrogen, lyophilized, placed in a 2 mL Eppendorf tube with 4 stainless steel 0.5 mm spheres and milled with a mixer mill for 60 s at 30 Hz.
Total RNA was extracted with RNAzol ®RT (Sigma-Aldrich, St. Louis, MO, USA) from 20–30 μg of lyophilized ground plant tissue, following the manufacturer’s instructions. The integrity of RNA was evaluated by denaturing gel electrophoresis. Concentration and purity were assessed by measuring OD260/OD230 and OD260/OD280 absorbance ratios using a microplate spectrophotometer (Epoch, Bio-Tek, Winooski, VT, USA). First-strand cDNA was synthetized from 1 μg of total RNA using Fast Gene® Scriptase II (Nippon Genetics Europe, Düren, Germany) and oligo dT, following the manufacturer’s instructions. Primers used were published in previous studies (Table S1) with the exception of ICS2, which was designed using Primer3 v.4.1.0) [50] with B. napus specific gene sequences from the Genbank (https://www.ncbi.nlm.nih.gov/genbank/ (accessed on 21 November 2021)) data base.
Gene expression was measured by qPCR with the equipment described in 2.6. The reaction mixture contained the equivalent of 5 ng total RNA, 5 μL of 2× qPCRBIO SyGreen Low-ROX (PCRBIOSYSTEMS, London, UK), 0.2 μL of 10 μM of each primer and 3.6 μL of 1 μL of DNA template solution, completed to a total of 10 μL of final volume. The program consisted of 95 °C for 2 min, 40 cycles of 95 °C for 10 s and 60 °C for 30 s. Amplicon specificity was controlled by melting curve analysis as previously described. The relative expression of each gene was calculated using the 2−∆∆CT method, with correction for primer efficiency, tested using a five-dilution series of the template [51], normalized to the endogenous reference gene ACTIN and subsequently normalized to those in the control plants. For this study, we selected the following genes in the hormonal and glucosinolate (GSL) pathway: abscisic acid (ABA) biosynthesis: ABA2 (xanthoxin dehydrogenase); ethylene (ET) biosynthesis: ACO (1-aminocyclopropane-1-carboxylic acid oxidase); ET downstream signaling: ERF2 (ethylene-responsive transcription factor 2); salicylic acid (SA) synthesis: PAL (phenylalanine ammonia-lyase) and ICS2 (isochorismate synthase 2); SA downstream signaling: PR1 (pathogenesis-related protein 1); jasmonic acid (JA) synthesis: AOS (allene oxide synthase); JA signaling: PDF 1.2 (defensin-like protein 16); aliphatic GSL biosynthesis: CYP83A1 (cytochrome P450 83A1) and indole GSLs biosynthesis: CYP79B2 (cytochrome P450 79B2).
2.8. Data Analysis
Data exploration and statistical analyses were performed with the software R 4.0.3 [52]. DNA, CFU and gene expression data were log transformed, and plant weight was transformed to the power of two to meet the assumptions of normality and homogeneity of variance and analyzed with one-way ANOVA. Significant differences between treatments were evaluated post hoc with the Fisher LSD test (Library agricolae, [53]). AUDPC values were analyzed with a generalized linear model (GLM) with the quasi-Poisson family due to overdispersion.
3. Results
3.1. Direct and Systemic Effects of M. brunneum on V. longisporum Plant Colonization and Disease Development
Metarhizium brunneum inhibited in vitro colony growth of V. longisporum when grown in dual culture. The inhibition was already evident after 14 d, when the fungal pathogen area was 13% smaller in dual culture than in single culture. By day 23, the growth inhibition was 42% (Table S1, Figure 1a). Vice versa, V. longisporum did not affect M. brunneum growth when grown in dual culture. The colony area of M. brunneum was marginally higher at 17 d in the dual culture (p = 0.091; F1,8 = 3.68; Table S1). Increased mycelia production (17 d), and localized spore formation (23 d) were observed on the side confronting the V. longisporum colony (Figure 1b).
3.2. Effect of Co-Inoculation M. brunneum on V. longisporum Disease Development
Co-inoculation with M. brunneum reduced the severity of symptoms of Verticillium stem striping disease. Although the shoot biomass of plants co-inoculated with both fungi was lower than that of control plants at 28 dpi, plants inoculated with both fungi had higher root dry weight (Figure 2b) and larger leaf area (Figure 2c) than plants inoculated with V. longisporum alone. The lower disease severity in plants co-inoculated with both fungi was reflected in the disease score which was lower at 21 and 28 dpi (Figure 2d). Furthermore, there was a delay in the colonization of the hypocotyl by the fungal pathogen in plants co-inoculated with M. brunneum. Specifically, the concentration of V. longisporum DNA increased between 21 and 28 dpi in plants inoculated with the fungal pathogen alone, while plants co-inoculated with both fungi had a lower amount of V. longisporum DNA at 28 dpi (Student’s t-test, t = −2.35, df = 7.24, p = 0.025).
However, the protective effects of M. brunneum co-inoculation were no longer observed at 35 dpi, where there were no significant differences in plant growth parameters, disease score or pathogen DNA in the hypocotyl between plants inoculated with V. longisporum alone and those co-inoculated with both fungi. We also measured M. brunneum in the hypocotyl tissues, which was detected only at 21 dpi (Figure 2f), with more plants showing endophytic growth in the dual inoculation treatment (three out of five plants) than in the treatment with only M. brunneum (one out of five).
3.3. Direct and Systemic Effects of M. brunneum on V. longisporum Plant Colonization and Disease Development
The split-root experiment revealed both local and systemic effects of M. brunneum inoculation on V. longisporum DNA levels in the roots and hypocotyl, as well as on plant biomass and disease progression.
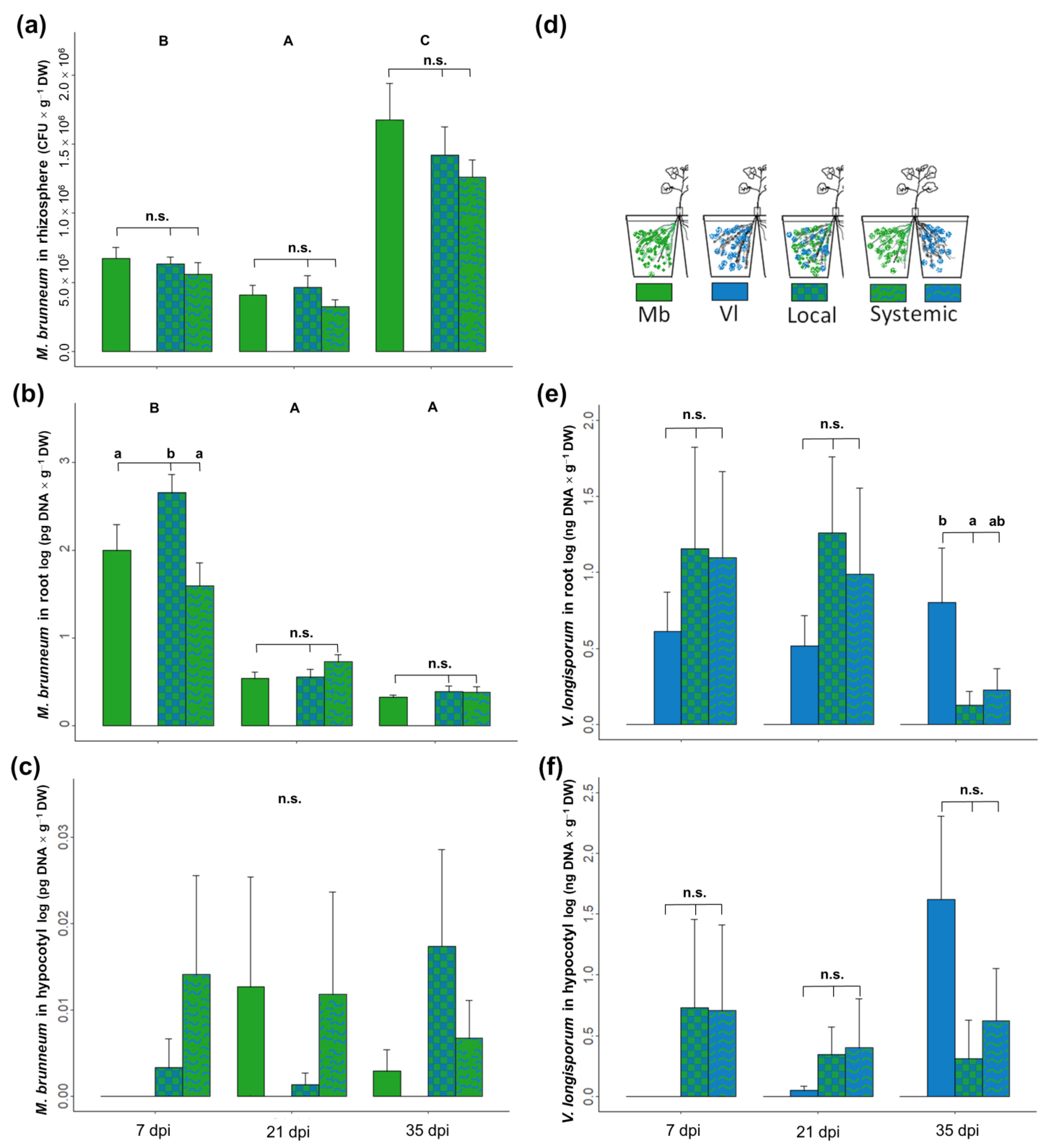
Both fungi were detected only in the part of the root where they were inoculated. The biomass of M. brunneum in the rhizosphere, measured as the number of colony forming units (CFUs), was not affected by presence of V. longisporum. However, M. brunneum CFUs changed over time, decreasing from 7 to 21 dpi and increasing at 35 dpi (Figure 3a). The amount of M. brunneum DNA in the roots was found to be significantly higher in the local compartment where V. longisporum was present, compared to roots where the entomopathogen was alone or in the systemic compartment at 7 dpi (Figure 3b).
DNA of M. brunneum in the hypocotyl was low and detected in few plants; it was detected in one plant per treatment at seven and 21 dpi and in about 30% of the plants at 35 dpi. There were no significant differences between treatments, but there was a tendency for a higher amount of DNA in the local treatment at 35 dpi (Figure 3c).
Verticillium longisporum concentration in plant roots was highly variable during the first three weeks (7 and 21 dpi) and was not affected by the presence of M. brunneum. However, pathogen DNA was lower in the local compartment at 35 dpi (Figure 3e, p = 0.037) and lower, though not significantly, in the systemic compartment (p = 0.072). Pathogen DNA in the hypocotyl (Figure 3f) was detected at 7 dpi, but only in one (Local) or two (Systemic) plants. At 35 dpi, there was a lower amount of V. longisporum DNA in the local and systemic treatments, but differences were only marginally statistically significant (p = 0.061 and p = 0.092, respectively).
Inoculation with M. brunneum reduced plant stunting caused by V. longisporum infection as well as the AUDPC in both local and systemic treatments (Figure 4). We only observed effects on plant biomass at 35 dpi, where the V. longisporum treatment had lower biomass than the other treatments (Figure 4a, lm, F4,50 = 3.28, p = 0.018; Vl vs. Control p = 0.006). This was also observed for disease progression, where the V. longisporum treatment had a higher AUDPC (Figure 4b, lm, F4,50 = 3.62, p = 0.002).
3.4. Direct and Systemic Plant Responses to V. longisporum and M. brunneum: Gene Expression
Inoculation with M. brunneum alone affected the plant’s SA pathway by upregulating the SA downstream signaling gene PR1, but not the SA biosynthesis genes PAL and ICS, compared to the control. Likewise, the JA/ET downstream signaling gene PDF1.2 was upregulated in the local and systemic root compartments (Figure 5d). Metarhizium brunneum inoculation also induced changes in the ET pathway, upregulating ET synthesis (ACO) in the local compartment (Figure 5e) and decreasing the downstream signaling gene ERF both in the local and systemic compartment. The plant responded to V. longisporum infection by induction of the SA-responsive gene PR1 (Figure 5c). The JA/ET downstream marker PDF1.2 was also induced in the roots in both local (Vl-L) and systemic (Vl-S) compartments (Figure 5d). Interestingly, there was only a systemic but not a local plant response to the pathogen in the ET biosynthesis pathway with regard to ACO induction (Figure 5e), while ERF was slightly downregulated when compared with control roots (Figure 5f). There were no changes in the expression of genes involved in the biosynthesis of JA (AOS, Figure S1), abscisic acid (ABA2, Figure 5g) or glucosinolates (GSL) (CYP83A1 and CYP79B2, Figure 5h,i).
Interestingly, when M. brunneum was applied together in the same root compartment as V. longisporum (Mb-L/VI-L), roots had significantly higher PAL expression when compared with the control (Figure 5a). The induction of PR1 in V. longisporum infected roots increased when M. brunneum was present in the systemic compartment (Vl-L/Mb-S), compared to the treatment with the pathogen alone (VL-L, Figure 5c). In the local treatment (Vl-L/Mb-L), induction of PR1 was intermediate between the pathogen-only treatment and the systemic treatment and did not significantly differ from them. There was no local or systemic inhibition of the ERF gene (Figure 5f) when both fungi were colonizing the plant. The abscisic acid biosynthesis gene ABA2 was induced in the compartments where M. brunneum was present (Mb-L/Vl-L and Mb-L/Vl-S, Figure 5g). In the systemic treatment with M. brunneum and V. longisporum in separate root compartments, significant downregulation of the aliphatic GSL biosynthesis gene CYP83A1 was observed in the Vl compartment (VI-L/Mb-S, Figure 5h), while for the indol GSL biosynthesis gene CYP79B2 downregulation was found in the Mb compartment (VI-L/Mb-S, Figure 5i).
4. Discussion
This study examined how M. brunneum, a soil-borne fungal entomopathogen, protects oilseed rape plants from V. longisporum, a soil-borne pathogen. The results show that M. brunneum effectively delays the colonization of plant roots by V. longisporum, leading to a significant reduction in disease symptoms. The research highlights the importance of direct competition and antibiosis in this process, as M. brunneum thrives in the presence of the pathogen while inhibiting its growth. Additionally, using a split-root setup, we observed that M. brunneum has a notable impact on the root’s defense response both locally and systemically by priming the SA pathway.
The in vitro confrontation assays conducted in this study demonstrated that M. brunneum inhibited the growth of V. longisporum by forming an inhibition halo. This inhibition may be attributed to the production of secondary metabolites with antibiotic effects [27]. Fungal inhibition in dual cultures with Metarhizium species was observed against a variety of other plant pathogens [20,21,22,54]. Crude extracts and partially purified fractions of Metarhizium have also been reported to inhibit fungal pathogens in previous studies [20,54,55,56]. Moreover, M. brunneum volatile organic compounds (VOCs) also inhibit plant pathogenic fungi [57]. Interestingly, our results indicate that the presence of the pathogen V. longisporum did not hinder the growth of M. brunneum in the confrontation assay. In fact, there was a slight inclination towards increased colony growth. Moreover, in pot experiments, the presence of V. longisporum did not affect the growth of M. brunneum in the soil. On the contrary, the entomopathogen was more abundant in the roots when V. longisporum was present. These findings suggest that the presence of other fungal mycelia may stimulate the growth of M. brunneum, leading to increased colonization of the root and potentially slowing down the spread of the pathogen’s infection.
After penetrating the roots, V. longisporum colonizes the xylem vessels and grows into the shoot, where it stays restricted to single vessels until later, when the fungus starts its saprophytic phase by invading the stem parenchyma and producing microsclerotia in the shoot tissues [41,58]. In a previous study by Eynck et al. [41], it was found that V. longisporum is initially detected at low levels in B. napus plants within the first four weeks after inoculation. However, its presence sharply increases in the hypocotyl at 35 dpi, as determined by quantifying the amount of its DNA. We aimed to investigate the impact of M. brunneum on the colonization of V. longisporum in the root and hypocotyl. Our observations revealed that at the root level (measured only in the split-root setup), the abundance of the pathogen was significantly lower at 35 dpi in root compartments where M. brunneum was present. In addition, during the co-inoculation experiment, we noticed that the abundance of V. longisporum in the hypocotyl was lower at 28 dpi. These findings suggest that while there were no apparent signs of V. longisporum inhibition in the roots at 7 and 21 dpi, a competitive interaction in the root zone resulted in reduced colonization of both the roots and hypocotyls by the pathogen. As a consequence, we observed diminished disease symptoms and improved plant growth in both the co-inoculation and split-root experiments. These results indicate that the presence of M. brunneum influenced the colonization dynamics of V. longisporum. In a previous study, it was found that pre-inoculation with the endophyte V. isaacii Vt305 also led to reduced plant colonization and symptom development caused by V. longisporum [59]. Although the specific mechanism was not investigated, the authors proposed that induced resistance could be responsible for this observed effect.
Our results from the split-root experiment further support the potential involvement of induced resistance in addition to the direct control mechanisms discussed earlier. Interestingly, even when M. brunneum was inoculated in a separate compartment from the pathogen, a protective effect was observed in the systemic treatment. In this case, the AUDPC values were lower, and there was no significant decrease in plant biomass compared to the treatment with the pathogen alone. Additionally, there was a marginal decrease in the amount of pathogen DNA detected in both the roots and hypocotyls.
Beneficial root-associated fungi can elicit induced systemic responses (ISR) in plants and prime them to respond more effectively against pathogens [4,31]. The impact of Metarhizium species on plant defenses is not consistent, as studies have shown contradictory results. For example, M. anisopliae M202-1 was found to suppress plant defenses in peanut roots [32], while different species of Metarhizium induced the SA and JA pathways in other plants [12,35,36]. In a study conducted previously, we observed no significant plant responses after five weeks of M. brunneum Gd12 inoculation [37]. However, our present study suggests that M. brunneum CB15III induces the upregulation of two key pathways: the SA pathway, indicated by an increased transcription of the PR1 gene, and the JA branch, resulting in upregulation of PDF1.2. In contrast, we observed the downregulation of ERF2, which is a protein involved in the transcriptional regulation of defense genes in response to ET and/or elicitors [60]. The inhibition of ET response has been observed both in M. anisopliae and the pathogen F. oxysporum [32]. This downregulation could potentially be a common response to root colonizers in plants.
ABA, SA and JA/ET pathways play a crucial role in the response of oilseed rape plants to V. longisporum, as demonstrated by comparative transcriptomic analysis [48]. Infection by V. longisporum results in the suppression of genes involved in ABA biosynthesis, induction of the SA signaling pathway and decreased response of the JA/ET pathway.
Another study investigated the phenylpropanoid and SA pathways in a resistant and susceptible variety of B. napus, demonstrating that the resistant line exhibited increased SA levels and elevated expression of SA marker genes PR1 and PR2 [49]. SA signaling activation was further supported by a study showing a two-fold increase in SA-activated PR1 expression in A. thaliana. Our findings align with these results, as we observed higher PR1 expression in both the local and systemic compartments of V. longisporum-inoculated plants. Furthermore, this study provides additional evidence supporting the role of the JA/ET signaling pathway in the plant response to V. longisporum. We found upregulation of the JA/ET downstream signaling gene PDF1.2 at both the local and systemic levels. A previous study reported the activation of PDF1.2 in oilseed rape [61] and A. thaliana but not in B. napus plants at 6 dpi [48]. Interestingly, we observed changes in ET markers exclusively in the systemic compartment. While the induction of ACO suggests activation of ET synthesis, the downregulation of ERF indicates inhibition of downstream signaling. Behrens et al. [48] also reported the induction of ethylene receptor 2 (ETR2) in B. napus at 3 dpi and A. thaliana at 6 dpi. The distinct systemic response of PDF1.2, ACO and ERF to V. longisporum inoculation suggests that the pathogen may locally manipulate JA/ET signaling while the plant is capable of activating systemic signaling.
Root associated beneficial fungi can modify the plant response to V. longisporum. For example, Trichoderma harzianum and Bacillus velezenis induced a priming response on B. napus against this pathogen that involved the activation of the JA and ET hormonal pathways [61]. Our observations suggest that M. brunneum altered the root response to V. longisporum both locally and systemically. Phenylalanine ammonia-lyase (PAL) is the first enzyme in the phenylpropanoid pathway, which is directly linked to increased resistance to V. longisporum [49]. We found that M. brunneum co-inoculation with the pathogen increased PAL gene expression. This is consistent with other studies showing that beneficial microorganisms can induce PAL gene expression or enzyme activity in response to a pathogen. For example, P. fluorescens, different strains of B. bassiana and T. asperellum T-203 induced PAL expression or activity in olive, tomato and cucumber plants infected with V. dahliae, Rhizoctonia solani and Pseudomona syringae, respectively [62,63,64].
Furthermore, we found that the gene expression of PR1 was higher in Verticillium-infected roots when the entomopathogenic fungus was present in the adjacent compartment. This suggests that the plant mounts a stronger response to the pathogen through the SA pathway, which leads to a systemic induction of plant defenses. Although priming of plants against pathogens through the SA pathway has been reported for other beneficial endophytes, our study is the first to report priming in response to Metarhizium inoculation.
It is known that during the early stages of infection, V. longisporum requires the suppression of ABA biosynthesis to establish itself [48]. However, in our study we did not observe any downregulation of ABA2 by V. longisporum. Instead, we found that M. brunneum induced an upregulation of this gene both locally and systemically when the pathogen was present. This finding suggests that the presence of M. brunneum may interfere with the pathogen’s manipulation of ABA biosynthesis, which could lead to increased resistance in the plant.
Glucosinolates (GSL) are secondary metabolites found in Brassica plants that serve as a defense mechanism [65] by exhibiting antimicrobial properties [66,67]. These compounds appear to be involved in the plant’s response to V. longisporum. For instance, the gene CYP79B2, that encodes for an enzyme involved in indolic GSL biosynthesis, is induced in A. thaliana plants when infected with the pathogen. Furthermore, the double mutant cyp79b2 cyp79b3 showed greater susceptibility to this pathogen [68,69]. GSLs also play a role in endophyte-plant interactions, as the root endophyte Piriformospora indica DSM 11827 induces the CYP79B2 gene. In its absence, as found in mutant cyp79B2 cyp79B3, the endophyte grows uncontrollably [70]. In our study, we found no evidence of CYP79B2 or CYP83A1 induction by V. longisporum in B. napus plants. Instead, we observed inhibition of CYP83A1 in the compartment inoculated with the pathogen when M. brunneum was present in the adjacent compartment and inhibition of CYP79B2 in the compartment inoculated with M. brunneum. These results suggest a possible synergistic inhibition of GSL biosynthesis by the two microorganisms. However, further investigations are needed to confirm this hypothesis.
Beneficial fungi can also induce plant resistance through the release of various metabolites in the zone of interaction [31,71]. These metabolites can be recognized as microbe-associated molecular patterns (MAMPs) by the plant immune system and trigger defense responses [31,72]. The mechanisms behind plant recognition of endophytic entomopathogenic fungi (EEF) and the induction of systemic resistance are not yet fully understood [7], but recent research suggests that the EEF B. bassiana (BG11 and FRh2) may elicit the upregulation of pattern recognition receptors in A. thaliana plants, indicating a potential for microbe-associated molecular pattern-triggered immunity [7,33]. There is growing evidence that EEF induces systemic resistance in the plant, both against insects and plant pathogens [33,36,37,38]. An important step in the future is to understand how these defense responses are elicited and to find MAMPs or other compounds responsible for the induction of plant defenses.
5. Conclusions
This study shows that M. brunneum delayed V. longisporum oilseed rape root colonization, resulting in decreased disease symptoms. The mechanisms involved include a faster colonization of M. brunneum, whose growth was stimulated by the pathogen’s presence. This has likely resulted in preempting the space and a competition for resources, together with antibiosis. Moreover, M. brunneum changed the plant’s response to the pathogen by locally activating PAL and ABA genes, suggesting an activation of the phenylpropanoid and abscisic acid pathways. Furthermore, with the split-root experiment we were able to prove that there is a plant-mediated effect, seen by improved plant growth and decreased disease symptoms when M. brunneum was in the systemic compartment. In addition, enhanced systemic induction of PR1 suggested a priming effect. So far, several studies have demonstrated the induction of systemic resistance by EEF. To our knowledge, we provide the first evidence of ISR by M. brunneum against a soil-borne pathogen.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof9080796/s1, Figure S1: Expression of allene oxide synthase AOS gene; Table S1: Primers used for qPCR amplification; Table S2: Summary statistics of fungal confrontation assays.
Author Contributions
Conceptualization, M.R., S.V. and C.P.-V.; methodology, M.R. and C.P.-V.; formal analysis, C.P.-V.; investigation, C.P.-V.; resources, M.R. and S.V.; data curation, C.P.-V.; writing—original draft preparation, C.P.-V.; writing—review and editing, M.R. and S.V.; visualization, C.P.-V.; supervision, M.R. and S.V.; project administration, M.R.; funding acquisition, M.R. and S.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding. C.P.-V. was supported in part by a doctoral studies scholarship from the “Ministerio de Ciencia, Tecnología e Innovación” -COLCIENCIAS-.
Data Availability Statement
Data will be available upon request.
Acknowledgments
The authors acknowledge the support by the Open Access Publication Funds of the Goettingen University.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Torsvik, V.; Øvreås, L. Microbial Diversity and Function in Soil: From Genes to Ecosystems. Curr. Opin. Microbiol. 2002, 5, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Bakker, P.A.H.M.; Berendsen, R.L.; Doornbos, R.F.; Wintermans, P.C.A.; Pieterse, C.M.J. The Rhizosphere Revisited: Root Microbiomics. Front. Plant Sci. 2013, 4, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, A.; Schmid, M.; van Tuinen, D.; Berg, G. Plant-Driven Selection of Microbes. Plant Soil 2009, 321, 235–257. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced Systemic Resistance by Beneficial Microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The Rhizosphere Microbiome and Plant Health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Bruck, D.J. Ecology of Metarhizium anisopliae in Soilless Potting Media and the Rhizosphere: Implications for Pest Management. Biol. Control 2005, 32, 155–163. [Google Scholar] [CrossRef]
- Hu, S.; Bidochka, M.J. Root Colonization by Endophytic Insect-Pathogenic Fungi. J. Appl. Microbiol. 2021, 130, 570–581. [Google Scholar] [CrossRef]
- Sasan, R.K.; Bidochka, M.J. The Insect-Pathogenic Fungus Metarhizium robertsii (Clavicipitaceae) Is Also an Endophyte That Stimulates Plant Root Development. Am. J. Bot. 2012, 99, 101–107. [Google Scholar] [CrossRef]
- Behie, S.W.; Jones, S.J.; Bidochka, M.J. Plant Tissue Localization of the Endophytic Insect Pathogenic Fungi Metarhizium and Beauveria. Fungal Ecol. 2015, 13, 112–119. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Khan, S.A.; Kang, S.M.; Shinwari, Z.K.; Kamran, M.; ur Rehman, S.; Kim, J.G.; Lee, I.J. Pure Culture of Metarhizium anisopliae LHL07 Reprograms Soybean to Higher Growth and Mitigates Salt Stress. World J. Microbiol. Biotechnol. 2012, 28, 1483–1494. [Google Scholar] [CrossRef]
- Verma, P.; Khan, S.A.; Mathur, A.K.; Shanker, K.; Kalra, A. Fungal Endophytes Enhanced the Growth and Production Kinetics of Vinca minor Hairy Roots and Cell Suspensions Grown in Bioreactor. Plant Cell Tissue Organ Cult. 2014, 118, 257–268. [Google Scholar] [CrossRef]
- Ahmad, I.; del Mar Jiménez-Gasco, M.; Luthe, D.S.; Shakeel, S.N.; Barbercheck, M.E. Endophytic Metarhizium robertsii Promotes Maize Growth, Suppresses Insect Growth, and Alters Plant Defense Gene Expression. Biol. Control 2020, 144, 104167. [Google Scholar] [CrossRef]
- Jaber, L.R. Seed Inoculation with Endophytic Fungal Entomopathogens Promotes Plant Growth and Reduces Crown and Root Rot (CRR) Caused by Fusarium culmorum in Wheat. Planta 2018, 248, 1525–1535. [Google Scholar] [CrossRef]
- Jaber, L.R.; Enkerli, J. Fungal Entomopathogens as Endophytes: Can They Promote Plant Growth? Biocontrol Sci. Technol. 2017, 27, 28–41. [Google Scholar] [CrossRef]
- Sanchez-Rodriguez, A.R.; Barron, V.; Del Campillo, M.C.; Quesada-Moraga, E. The Entomopathogenic Fungus Metarhizium brunneum: A Tool for Alleviating Fe Chlorosis. Plant Soil 2016, 406, 295–310. [Google Scholar] [CrossRef]
- Behie, S.W.; Zelisko, P.M.; Bidochka, M.J. Endophytic Insect-Parasitic Fungi Translocate Nitrogen Directly from Insects to Plants. Science 2012, 336, 1576–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krell, V.; Unger, S.; Jakobs-Schoenwandt, D.; Patel, A.V. Importance of Phosphorus Supply through Endophytic Metarhizium brunneum for Root:Shoot Allocation and Root Architecture in Potato Plants. Plant Soil 2018, 430, 87–97. [Google Scholar] [CrossRef]
- Gange, A.C.; Koricheva, J.; Currie, A.F.; Jaber, L.R.; Vidal, S. Meta-Analysis of the Role of Entomopathogenic and Unspecialized Fungal Endophytes as Plant Bodyguards. New Phytol. 2019, 223, 2002–2010. [Google Scholar] [CrossRef] [Green Version]
- Vega, F.E. The Use of Fungal Entomopathogens as Endophytes in Biological Control: A Review. Mycologia 2018, 110, 4–30. [Google Scholar] [CrossRef]
- Sasan, R.K.; Bidochka, M.J. Antagonism of the Endophytic Insect Pathogenic Fungus Metarhizium robertsii against the Bean Plant Pathogen Fusarium solani f. Sp. Phaseoli. Can. J. Plant Pathol. 2013, 35, 288–293. [Google Scholar] [CrossRef]
- Miranda-Fuentes, P.; García-Carneros, A.B.; Montilla-Carmona, A.M.; Molinero-Ruiz, L. Evidence of Soil-located Competition as the Cause of the Reduction of Sunflower Verticillium Wilt by Entomopathogenic Fungi. Plant Pathol. 2020, 69, 1492–1503. [Google Scholar] [CrossRef]
- Jaber, L.R.; Alananbeh, K.M. Fungal Entomopathogens as Endophytes Reduce Several Species of Fusarium Causing Crown and Root Rot in Sweet Pepper (Capsicum annuum L.). Biol. Control 2018, 126, 117–126. [Google Scholar] [CrossRef]
- Rivas-Franco, F.; Hampton, J.G.; Morán-Diez, M.E.; Narciso, J.; Rostás, M.; Wessman, P.; Jackson, T.A.; Glare, T.R. Effect of Coating Maize Seed with Entomopathogenic Fungi on Plant Growth and Resistance against Fusarium graminearum and Costelytra giveni. Biocontrol Sci. Technol. 2019, 29, 877–900. [Google Scholar] [CrossRef]
- Gupta, R.; Keppanan, R.; Leibman-Markus, M.; Rav-David, D.; Elad, Y.; Ment, D.; Bar, M. The Entomopathogenic Fungi Metarhizium brunneum and Beauveria bassiana Promote Systemic Immunity and Confer Resistance to a Broad Range of Pests and Pathogens in Tomato. Phytopathology 2022, 112, 784–793. [Google Scholar] [CrossRef] [PubMed]
- St. Leger, R.J.; Wang, J.B. Metarhizium: Jack of All Trades, Master of Many. Open Biol. 2020, 10, 200307. [Google Scholar] [CrossRef]
- Ownley, B.H.; Gwinn, K.D.; Vega, F.E. Endophytic Fungal Entomopathogens with Activity against Plant Pathogens: Ecology and Evolution. BioControl 2010, 55, 113–128. [Google Scholar] [CrossRef]
- Moonjely, S.; Barelli, L.; Bidochka, M.J. Insect Pathogenic Fungi as Endophytes. Adv. Genet. 2016, 94, 107–135. [Google Scholar] [CrossRef]
- Jaber, L.R.; Ownley, B.H. Can We Use Entomopathogenic Fungi as Endophytes for Dual Biological Control of Insect Pests and Plant Pathogens? Biol. Control 2018, 116, 36–45. [Google Scholar] [CrossRef]
- Dreischhoff, S.; Das, I.S.; Jakobi, M.; Kasper, K.; Polle, A. Local Responses and Systemic Induced Resistance Mediated by Ectomycorrhizal Fungi. Front. Plant Sci. 2020, 11, 590063. [Google Scholar] [CrossRef]
- Hill, E.M.; Robinson, L.A.; Abdul-sada, A.; Vanbergen, A.J.; Hodge, A.; Hartley, S.E.; Hartley, S.E. Arbuscular Mycorrhizal Fungi and Plant Chemical Defence: Effects of Colonisation on Aboveground and Belowground Metabolomes. J. Chem. Ecol. 2018, 44, 198–208. [Google Scholar] [CrossRef] [Green Version]
- Shoresh, M.; Harman, G.E.; Mastouri, F. Induced Systemic Resistance and Plant Responses to Fungal Biocontrol Agents. Annu. Rev. Phytopathol. 2010, 48, 21–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, K.; Wang, F.; Nong, X.; McNeill, M.R.; Liu, S.; Wang, G.; Cao, G.; Zhang, Z. Response of Peanut Arachis hypogaea Roots to the Presence of Beneficial and Pathogenic Fungi by Transcriptome Analysis. Sci. Rep. 2017, 7, 964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raad, M.; Glare, T.R.; Brochero, H.L.; Müller, C.; Rostás, M. Transcriptional Reprogramming of Arabidopsis thaliana Defence Pathways by the Entomopathogen Beauveria bassiana Correlates with Resistance Against a Fungal Pathogen but Not Against Insects. Front. Microbiol. 2019, 10, 615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, X.; Zhao, X.; Huang, S.; Deng, J.; Li, X.; Luo, Z.; Zhang, Y. Pest Management via Endophytic Colonization of Tobacco Seedlings by the Insect Fungal Pathogen Beauveria bassiana. Pest Manag. Sci. 2021, 77, 2007–2018. [Google Scholar] [CrossRef]
- Cotes, B.; Thöming, G.; Amaya-Gómez, C.V.; Novák, O.; Nansen, C. Root-Associated Entomopathogenic Fungi Manipulate Host Plants to Attract Herbivorous Insects. Sci. Rep. 2020, 10, 22424. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Franco, F.; Hampton, J.G.; Narciso, J.; Rostás, M.; Wessman, P.; Saville, D.J.; Jackson, T.A.; Glare, T.R. Effects of a Maize Root Pest and Fungal Pathogen on Entomopathogenic Fungal Rhizosphere Colonization, Endophytism and Induction of Plant Hormones. Biol. Control 2020, 150, 104347. [Google Scholar] [CrossRef]
- Posada-Vergara, C.; Lohaus, K.; Alhussein, M.; Vidal, S.; Rostás, M. Root Colonization by Fungal Entomopathogen Systemically Primes Belowground Plant Defense against Cabbage Root Fly. J. Fungi 2022, 8, 969. [Google Scholar] [CrossRef]
- Cachapa, J.C.; Meyling, N.V.; Burow, M.; Hauser, T.P. Induction and Priming of Plant Defense by Root-Associated Insect-Pathogenic Fungi. J. Chem. Ecol. 2020, 47, 112–122. [Google Scholar] [CrossRef]
- Qing, Y.; Ourry, M.; Burow, M.; Meyling, N.V.; Hauser, T.P. Defense Priming in Cabbage (Brassica oleracea) by Insect-Pathogenic Fungi. Arthropod Plant Interact. 2023, 17, 275–287. [Google Scholar] [CrossRef]
- Depotter, J.R.L.L.; Deketelaere, S.; Inderbitzin, P.; Von Tiedemann, A.; Höfte, M.; Subbarao, K.V.; Wood, T.A.; Thomma, B.P.H.J. Verticillium longisporum, the Invisible Threat to Oilseed Rape and Other Brassicaceous Plant Hosts. Mol. Plant Pathol. 2016, 17, 1004–1016. [Google Scholar] [CrossRef] [Green Version]
- Eynck, C.; Koopmann, B.; Grunewaldt-Stoecker, G.; Karlovsky, P.; von Tiedemann, A. Differential Interactions of Verticillium longisporum and V. Dahliae with Brassica napus Detected with Molecular and Histological Techniques. Eur. J. Plant Pathol. 2007, 118, 259–274. [Google Scholar] [CrossRef]
- Eynck, C.; Koopmann, B.; Karlovsky, P.; Von Tiedemann, A. Internal Resistance in Winter Oilseed Rape Inhibits Systemic Spread of the Vascular Pathogen Verticillium longisporum. Phytopathology 2009, 99, 802–811. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madden, L.V.; Hughes, G.; Bosch, F. The Study of Plant Disease Epidemics; The American Phytopathological Society: St. Paul, MI, USA, 2007. [Google Scholar]
- Strasser, H.; Forer, A.; Schinner, F. Development of Media for the Selective Isolation and Maintenance of Virulence of Beauveria brongniartii. In Proceedings of the 3rd International Workshop on Microbial Control of Soil Dwelling Pests, Lincoln, New Zealand, 21–23 February 1996; Jackson, T.A., Glare, T.A., Eds.; AgResearch Lincoln: Lincoln, New Zealand, 1996; pp. 125–130. [Google Scholar]
- Brandfass, C.; Karlovsky, P. Simultaneous Detection of Fusarium culmorum and F. graminearum in Plant Material by Duplex PCR with Melting Curve Analysis. BMC Microbiol. 2006, 6, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, S.; Rehner, S.A.; Widmer, F.; Enkerli, J. A PCR-Based Tool for Cultivation-Independent Detection and Quantification of Metarhizium Clade 1. J. Invertebr. Pathol. 2011, 108, 106–114. [Google Scholar] [CrossRef]
- Behrens, F.H.; Schenke, D.; Hossain, R.; Ye, W.; Schemmel, M.; Bergmann, T.; Häder, C.; Zhao, Y.; Ladewig, L.; Zhu, W.; et al. Suppression of Abscisic Acid Biosynthesis at the Early Infection Stage of Verticillium longisporum in Oilseed Rape (Brassica Napus). Mol. Plant Pathol. 2019, 20, 1645–1661. [Google Scholar] [CrossRef]
- Zheng, X.; Koopmann, B.; von Tiedemann, A. Role of Salicylic Acid and Components of the Phenylpropanoid Pathway in Basal and Cultivar-Related Resistance of Oilseed Rape (Brassica napus) to Verticillium longisporum. Plants 2019, 8, 491. [Google Scholar] [CrossRef] [Green Version]
- Koressaar, T.; Lepamets, M.; Kaplinski, L.; Raime, K.; Andreson, R.; Remm, M. Primer3_masker: Integrating Masking of Template Sequence with Primer Design Software. Bioinformatics 2018, 34, 1937–1938. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Mendiburu, F.; Yaseen, M. Agricolae: Statistical Procedures for Agricultural Research 2020. R Package Version 1.4.0. Available online: https://myaseen208.github.io/agricolae/https://cran.r-project.org/package=agricolae (accessed on 27 May 2021).
- Lozano-Tovar, M.D.; Ortiz-Urquiza, A.; Garrido-Jurado, I.; Trapero-Casas, A.; Quesada-Moraga, E. Assessment of Entomopathogenic Fungi and Their Extracts against a Soil-Dwelling Pest and Soil-Borne Pathogens of Olive. Biol. Control 2013, 67, 409–420. [Google Scholar] [CrossRef]
- Guigón López, C.; Muñoz Castellanos, L.N.; Flores Ortiz, N.A.; González González, J.A. Control of Powdery Mildew (Leveillula taurica) Using Trichoderma asperellum and Metarhizium anisopliae in Different Pepper Types. BioControl 2019, 64, 77–89. [Google Scholar] [CrossRef]
- Lozano-Tovar, M.D.; Garrido-Jurado, I.; Quesada-Moraga, E.; Raya-Ortega, M.C.; Trapero-Casas, A. Metarhizium brunneum and Beauveria bassiana Release Secondary Metabolites with Antagonistic Activity against Verticillium dahliae and Phytophthora megasperma Olive Pathogens. Crop Prot. 2017, 100, 186–195. [Google Scholar] [CrossRef]
- Hummadi, E.H.; Cetin, Y.; Demirbek, M.; Kardar, N.M.; Khan, S.; Coates, C.J.; Eastwood, D.C.; Dudley, E.; Maffeis, T.; Loveridge, J.; et al. Antimicrobial Volatiles of the Insect Pathogen Metarhizium brunneum. J. Fungi 2022, 8, 326. [Google Scholar] [CrossRef] [PubMed]
- Knüfer, J.; Lopisso, D.T.; Koopmann, B.; Karlovsky, P.; von Tiedemann, A. Assessment of Latent Infection with Verticillium longisporum in Field-Grown Oilseed Rape by QPCR. Eur. J. Plant Pathol. 2017, 147, 819–831. [Google Scholar] [CrossRef]
- Tyvaert, L.; França, S.C.; Debode, J.; Höfte, M. The Endophyte Verticillium Vt305 Protects Cauliflower against Verticillium Wilt. J. Appl. Microbiol. 2014, 116, 1563–1571. [Google Scholar] [CrossRef]
- Nakano, T.; Nishiuchi, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Studies on Transcriptional Regulation of Endogenous Genes by ERF2 Transcription Factor in Tobacco Cells. Plant Cell Physiol. 2006, 47, 554–558. [Google Scholar] [CrossRef] [Green Version]
- Hafiz, F.B.; Moradtalab, N.; Goertz, S.; Rietz, S.; Dietel, K.; Rozhon, W.; Humbeck, K.; Geistlinger, J.; Neumann, G.; Schellenberg, I. Synergistic Effects of a Root-Endophytic Trichoderma Fungus and Bacillus on Early Root Colonization and Defense Activation Against Verticillium longisporum in Rapeseed. Mol. Plant-Microbe Interact. 2022, 35, 380–392. [Google Scholar] [CrossRef]
- Azadi, N.; Shirzad, A.; Mohammadi, H. A Study of Some Biocontrol Mechanisms of Beauveria bassiana against Rhizoctonia disease on Tomato. Acta Biol. Szeged. 2016, 60, 119–127. [Google Scholar]
- Cabanás, C.G.L.; Sesmero, R.; Valverde-Corredor, A.; Javier López-Escudero, F.; Mercado-Blanco, J. A Split-Root System to Assess Biocontrol Effectiveness and Defense-Related Genetic Responses in above-Ground Tissues during the Tripartite Interaction Verticillium dahliae-Olive-Pseudomonas fluorescens PICF7 in Roots. Plant Soil 2017, 417, 433–452. [Google Scholar] [CrossRef]
- Yedidia, I.; Shoresh, M.; Kerem, Z.; Benhamou, N.; Kapulnik, Y.; Chet, I. Concomitant Induction of Systemic Resistance to Pseudomonas syringae Pv. Lachrymans in Cucumber by Trichoderma asperellum (T-203) and Accumulation of Phytoalexins. Appl. Environ. Microbiol. 2003, 69, 7343–7353. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Wang, H.; Xie, J.; Lv, J.; Zhang, G.; Hu, L.; Luo, S.; Li, L.; Yu, J. The Roles of Cruciferae Glucosinolates in Disease and Pest Resistance. Plants 2021, 10, 1097. [Google Scholar] [CrossRef] [PubMed]
- Bednarek, P.; Piślewska-Bednarek, M.; Svatoš, A.; Schneider, B.; Doubský, J.; Mansurova, M.; Humphry, M.; Consonni, C.; Panstruga, R.; Sanchez-Vallet, A.; et al. A Glucosinolate Metabolism Pathway in Living Plant Cells Mediates Broad-Spectrum Antifungal Defense. Science 2009, 323, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Vig, A.P.; Rampal, G.; Thind, T.S.; Arora, S. Bio-Protective Effects of Glucosinolates—A Review. LWT Food Sci. Technol. 2009, 42, 1561–1572. [Google Scholar] [CrossRef]
- Fröschel, C. In-Depth Evaluation of Root Infection Systems Using the Vascular Fungus Verticillium longisporum as Soil-Borne Model Pathogen. Plant Methods 2021, 17, 57. [Google Scholar] [CrossRef] [PubMed]
- Iven, T.; König, S.; Singh, S.; Braus-Stromeyer, S.A.; Bischoff, M.; Tietze, L.F.; Braus, G.H.; Lipka, V.; Feussner, I.; Dröge-Laser, W. Transcriptional Activation and Production of Tryptophan-Derived Secondary Metabolites in Arabidopsis Roots Contributes to the Defense against the Fungal Vascular Pathogen Verticillium longisporum. Mol. Plant 2012, 5, 1389–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nongbri, P.L.; Johnson, J.M.; Sherameti, I.; Glawischnig, E.; Halkier, B.A.; Oelmüller, R.; Botany, G.; Physiology, P.; Jena, F.; Str, D. Indole-3-Acetaldoxime-Derived Compounds Restrict Root Colonization in the Beneficial Interaction Between Arabidopsis Roots and the Endophyte Piriformospora indica. Mol. Plant Microbe Interact. 2012, 25, 1186–1197. [Google Scholar] [CrossRef] [Green Version]
- Harman, G.E.; Petzoldt, R.; Comis, A.; Chen, J. Interactions between Trichoderma harzianum Strain T22 and Maize Inbred Line Mo17 and Effects of These Interactions on Diseases Caused by Pythiuin ultimum and Colletotrichum graminicola. Phytopathology 2004, 94, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Newman, M.A.; Sundelin, T.; Nielsen, J.T.; Erbs, G. MAMP (Microbe-Associated Molecular Pattern) Triggered Immunity in Plants. Front. Plant Sci. 2013, 4, 139. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
In vitro inhibition of Verticillium longisporum growth by Metarhizium brunneum. (a). Fungal colony area in single vs dual-culture plates; (b) detail of the confrontation zone. Fungal colonies grew in potato dextrose agar (PDA) media for 23 days either alone (single) or in confrontation with each other (dual). F1,8 = 44.8; *** p < 0.001, n = 4. (b) Close-up of the confrontation zone.
Figure 1.
In vitro inhibition of Verticillium longisporum growth by Metarhizium brunneum. (a). Fungal colony area in single vs dual-culture plates; (b) detail of the confrontation zone. Fungal colonies grew in potato dextrose agar (PDA) media for 23 days either alone (single) or in confrontation with each other (dual). F1,8 = 44.8; *** p < 0.001, n = 4. (b) Close-up of the confrontation zone.

Figure 2.
Effect of co-inoculation of oilseed rape roots with M. brunneum and V. longisporum on plant development and Verticillium disease progress. (a) Plant shoot dry weight; (b) Plant root dry weight; (c) Leaf area; (d) Disease score (see [41]; (e) V. longisporum DNA in hypocotyl; (f) M. brunneum DNA in hypocotyl. Plants were inoculated by root dipping 21 days after germination. For the dual inoculation (Mb/Vl), a mix of spores was used. Plants were harvested at 21, 28 and 35 days post-inoculation (dpi). Letters represent statistically significant differences within each date according to a linear model and Tukey’s post hoc test. * p < 0.05, Welch two sample t-test; n.s. = no significant differences. Bars represent means ± SE; n = 5.
Figure 2.
Effect of co-inoculation of oilseed rape roots with M. brunneum and V. longisporum on plant development and Verticillium disease progress. (a) Plant shoot dry weight; (b) Plant root dry weight; (c) Leaf area; (d) Disease score (see [41]; (e) V. longisporum DNA in hypocotyl; (f) M. brunneum DNA in hypocotyl. Plants were inoculated by root dipping 21 days after germination. For the dual inoculation (Mb/Vl), a mix of spores was used. Plants were harvested at 21, 28 and 35 days post-inoculation (dpi). Letters represent statistically significant differences within each date according to a linear model and Tukey’s post hoc test. * p < 0.05, Welch two sample t-test; n.s. = no significant differences. Bars represent means ± SE; n = 5.

Figure 3.
Colonization of rhizospheric soil (a), oilseed rape roots (b,e) and hypocotyls (c,f) by M. brunneum and/or V. longisporum. Plants grew in a split-root setup and were inoculated with fungal spores by root drenching. Treatment commenced with M. brunneum at transplanting (8 d) and was followed by V. longisporum after 7 d. Plants were harvested at 7, 21 and 35 dpi of V. longisporum. Panel (d) shows inoculation scheme and color patterns. Capital letters denote statistical differences between sampling dates; small letters denote significant differences among treatments within the same date. HDS Tukey; p < 0.05 according to a linear model; n.s. = no significant differences; bars represent means ± SE; n = 8.
Figure 3.
Colonization of rhizospheric soil (a), oilseed rape roots (b,e) and hypocotyls (c,f) by M. brunneum and/or V. longisporum. Plants grew in a split-root setup and were inoculated with fungal spores by root drenching. Treatment commenced with M. brunneum at transplanting (8 d) and was followed by V. longisporum after 7 d. Plants were harvested at 7, 21 and 35 dpi of V. longisporum. Panel (d) shows inoculation scheme and color patterns. Capital letters denote statistical differences between sampling dates; small letters denote significant differences among treatments within the same date. HDS Tukey; p < 0.05 according to a linear model; n.s. = no significant differences; bars represent means ± SE; n = 8.

Figure 4.
Shoot biomass (a) and area under the disease progress curve (AUDPC) (b) of oilseed rape plants inoculated with V. longisporum and/or M. brunneum. Plants grew in a split-root setup and were inoculated with fungal spores by root drenching. Treatment commenced with M. brunneum at transplanting (8 d) and was followed by V. longisporum after 7 d. Plants were harvested 35 d post-inoculation (dpi) of V. longisporum. Inoculation scheme of a given treatment is represented above each bar. Letters represent statistically significant differences within treatments according to a linear model and Tukey’s post hoc test. Bars represent means ± SE; n = 8.
Figure 4.
Shoot biomass (a) and area under the disease progress curve (AUDPC) (b) of oilseed rape plants inoculated with V. longisporum and/or M. brunneum. Plants grew in a split-root setup and were inoculated with fungal spores by root drenching. Treatment commenced with M. brunneum at transplanting (8 d) and was followed by V. longisporum after 7 d. Plants were harvested 35 d post-inoculation (dpi) of V. longisporum. Inoculation scheme of a given treatment is represented above each bar. Letters represent statistically significant differences within treatments according to a linear model and Tukey’s post hoc test. Bars represent means ± SE; n = 8.

Figure 5.
Expression of genes involved in defense signaling in roots of B. napus in response to V. longisporum and/or M. brunneum inoculation in a split-root setup. The left panel shows marker genes in the salicylic acid pathway: (a) PAL, phenylalanine ammonia lyase; (b) ICS, isochorismate synthase and (c) PR1, pathogenesis-related protein 1. The middle panel shows (d) marker gene in the jasmonic acid pathway PDF1.2 plant defensin 1.2, and for ethylene biosynthesis (e) ACO, 1-Aminocyclopropane-1-Carboxylic Acid Oxidase and downstream signaling (f) ERF2, ethylene response factor 2. The right panel displays genes involved in (g) abscisic acid biosynthesis ABA2, xanthoxin dehydrogenase; (h) aliphatic glucosinolate (GSL) CYP83A1, cytochrome P450 83A1 and (i) indol-GSL biosynthesis CYP79B2, cytochrome P450 79B2. The illustrations above the figures a, d and g show the fungal inoculation scheme, either with M. brunneum (Mb) or V. longisporum (Vl) in the local (-L) or adjacent (-S) compartment and either treatments in the same compartment (Mb-L/Vl-L) or in each adjacent compartment of the same plant (Vl-L/Mb-S; Mb-L/Vl-S). Fungi were inoculated by root dipping. Treatment commenced with M. brunneum at transplanting (8 d) and was followed by V. longisporum after 7 d. Plants were harvested 7 days after V. longisporum inoculation. Panel (j) shows inoculation scheme and color patterns. Different letters indicate significant differences among treatments (Fisher LSD, p < 0.05). Bars represent means ± SE; n = 4–7.
Figure 5.
Expression of genes involved in defense signaling in roots of B. napus in response to V. longisporum and/or M. brunneum inoculation in a split-root setup. The left panel shows marker genes in the salicylic acid pathway: (a) PAL, phenylalanine ammonia lyase; (b) ICS, isochorismate synthase and (c) PR1, pathogenesis-related protein 1. The middle panel shows (d) marker gene in the jasmonic acid pathway PDF1.2 plant defensin 1.2, and for ethylene biosynthesis (e) ACO, 1-Aminocyclopropane-1-Carboxylic Acid Oxidase and downstream signaling (f) ERF2, ethylene response factor 2. The right panel displays genes involved in (g) abscisic acid biosynthesis ABA2, xanthoxin dehydrogenase; (h) aliphatic glucosinolate (GSL) CYP83A1, cytochrome P450 83A1 and (i) indol-GSL biosynthesis CYP79B2, cytochrome P450 79B2. The illustrations above the figures a, d and g show the fungal inoculation scheme, either with M. brunneum (Mb) or V. longisporum (Vl) in the local (-L) or adjacent (-S) compartment and either treatments in the same compartment (Mb-L/Vl-L) or in each adjacent compartment of the same plant (Vl-L/Mb-S; Mb-L/Vl-S). Fungi were inoculated by root dipping. Treatment commenced with M. brunneum at transplanting (8 d) and was followed by V. longisporum after 7 d. Plants were harvested 7 days after V. longisporum inoculation. Panel (j) shows inoculation scheme and color patterns. Different letters indicate significant differences among treatments (Fisher LSD, p < 0.05). Bars represent means ± SE; n = 4–7.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Posada-Vergara, C.; Vidal, S.; Rostás, M. Local Competition and Enhanced Defense: How Metarhizium brunneum Inhibits Verticillium longisporum in Oilseed Rape Plants. J. Fungi 2023, 9, 796. https://doi.org/10.3390/jof9080796
AMA Style
Posada-Vergara C, Vidal S, Rostás M. Local Competition and Enhanced Defense: How Metarhizium brunneum Inhibits Verticillium longisporum in Oilseed Rape Plants. Journal of Fungi. 2023; 9(8):796. https://doi.org/10.3390/jof9080796
Chicago/Turabian StylePosada-Vergara, Catalina, Stefan Vidal, and Michael Rostás. 2023. "Local Competition and Enhanced Defense: How Metarhizium brunneum Inhibits Verticillium longisporum in Oilseed Rape Plants" Journal of Fungi 9, no. 8: 796. https://doi.org/10.3390/jof9080796
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.